
Эволюция в поджелудочной железе
Один из важных компонентов внутренней среды организма — глюкоза. Постоянство уровня ее содержания в крови обеспечивается, с одной стороны, инсулином (продуктом В-клеток панкреатических островков) — инсулярный компонент, а с другой — комплексом эндокринных клеток, обеспечивающих повышение уровня глюкозы (хромаффинная и интерреналовая ткань надпочечников, А-клетки панкреатических островков и некоторые другие эндокринные клетки), — контринсулярный компонент. С нарушением системы регуляции уровня глюкозы в крови связана эндокринная патология — сахарный диабет, гиперинсулинизм, опухоли эндокринной части поджелудочной железы и т. д. Чтобы понять сложившиеся в процессе исторического развития закономерности взаимоотношений инсулярного и контринсулярного аппаратов, рассмотрим формирование в ходе эволюции эндокринной части поджелудочной железы у позвоночных, а также факторы, определяющие их развитие.
Бесчерепные. Ланцетник по ряду морфологических и функциональных признаков близок к древнейшим предкам всех позвоночных животных. Именно этим и обусловлен непреходящий интерес к нему. У ланцетника отсутствует поджелудочная железа органного типа. Железистые клетки, гомологичные клеткам экзо- и эндокринной частей поджелудочной железы более высокоорганизованных позвоночных, содержатся в эпителии слизистой оболочки кишечника и в печоночном выросте. Известные английские исследователи Пирс и Ван Ноорден показали, что часть эндокринных клеток кишечника ланцетника содержит биогенные амины, другие клетки дают иммунофлюоресценцию на антисыворотки к инсулину, глюкагону и гастрину млекопитающих (рис. 11). Эти клетки содержат эндокринные секреторные гранулы. Таким образом, в слизистой кишечника ланцетника имеются клетки, гомологичные клеткам эндокринной части поджелудочной железы более высокоорганизованных позвоночных.

Локализация гормоноподобной активности в кишечнике ланцетника…
Круглоротые. У представителей круглоротых — миног и миксин, как и у ланцетника, поджелудочная железа органного типа отсутствует. Клетки, гомологичные экзокринным панкреацитам, формируются в эпителии слизистой или среди соединительной ткани, окружающей кишечник. Именно кишечный эпителий является эмбриональным источником островкового аппарата круглоротых. У личинки миноги (пескоройки) клетки кишечного эпителия мигрируют в собственный слой слизистой оболочки. Выселившиеся клетки образуют скопления эндокринных клеток. На этом этапе развития миноги отсутствует прямая связь эндокринных клеток с капиллярами, но начинается функционирование инсулярного аппарата. Секрет из эндокринных клеток выделяется по голокриновому способу и по межклеточным щелям выводится в капиллярное русло кишечника.
У взрослых особей продолжается миграция эндокринных клеток из кишечника крипт в стенку кишки. Одновременно с миграцией эндокринных клеток идет процесс васкуляризации инсулярного аппарата. Далее в центре группы клеток образуется полость и скопления клеток превращаются в фолликулы с диаметром 90—120 мкм — это так называемые фолликулы Лангерганса. Стенка фолликула образована одним слоем клеток, а полость заполнена коллоидоподобным веществом. Сотрудница кафедры зоологии Стокгольмского университета в Швеции Лайла Винблад отметила, что в осенне-зимний (речной) период жизни в инсулярной ткани миног и миксин преобладают скопления клеток, тогда как в преднерестовый (морской) период — фолликулы. На основании этих наблюдений автор предположила, что образование фолликулов предопределяется стабильными условиями обитания животных на дне глубоких морей, когда потребность животных в инсулине снижается, а избыток инсулинпродуцирующей ткани образует фолликулы.
Таким образом, у круглоротых инсулярный аппарат (рис. 12) представлен скоплениями эндокринных клеток в толще кишки, в окружающей ее соединительной ткани.или, наконец, в паренхиме печени. В определенные периоды функциональной деятельности скопления эндокринных клеток преобразуются в фолликулы Лангерганса. В составе фолликулов выявлены В-клетки, вырабатывающие инсулин, а также другой тип гранулированных клеток, функция которых не выяснена. Большинство современных исследователей склонно считать, что А-клетки в фолликулах Лангергансакруглоротых отсутствуют, однако в кишечнике имеются глюкагон-продуцирующие клетки (В 1976 г. известные американские исследователи Джек Бринн и Август Эппл (Университет Восточной Каролины и Университет Томаса Джефферсона, Филадельфия) обнаружили новые типы клеток в фолликулах круглоротых. Это открытие требует переоценки островкового органа круглоротых как продуцирующего только инсулин, поскольку обнаруженные клетки являются ацидофильными и дающими положительную реакцию на триптофан, что свойственно А-клеткам более высокоорганизованных позвоночных).

Схема гистогенеза инсулярного аппарата круглоротых…
Хрящевые рыбы. У хрящевых рыб (акулы, скаты, катранообразные) поджелудочная железа представлена уже компактным органом дольчатого строения, состоящим из экзо- и эндокринной частей. У всех изученных видов хрящевых рыб эндокринные клетки в поджелудочной железе располагаются по ходу средних и мелких выводных протоков, окружая их в виде эпителиального слоя — так называемый внешний эпителий. Кроме того, описаны и другие формы организации островко-вой ткани: скопления эндокринных клеток, прилежащие к стенке выводных протоков, а также свободно расположенные островки, утратившие связь с выводными протоками. Эти три формы сочетаются друг с другом с преобладанием тех или иных форм у различных видов хрящевых рыб. В отличие от круглоротых, у всех изученных видов хрящевых рыб в эндокринной части поджелудочной железы встречаются три типа гормонпродуцирующих клеток: А-, В- и D-клетки.
Таким образом, у хрящевых рыб поджелудочная железа формируется как самостоятельный орган; фолликулы Лангерганса заменяются внешним эпителием; в составе эндокринной части поджелудочной железы появляются А- и D-клетки. Известные советские ученые Ю. К. Елецкий и В. В. Яглов рассматривают внешний эпителий как исходную форму структурной организации эндокринной части поджелудочной железу, позвоночных (поскольку у многих видов хрящевых рыб в процессе органогенеза поджелудочной железы наблюдается миграция эндокринных клеток из внешнего эпителия в экзокринную ткань с формированием панкреатических островков, связанных и несвязанных с выводными протоками железы) и как первую эволюционную форму организации эндокринной части поджелудочной железы.
Костные рыбы. У кистеперой рыбы латимерии основная масса эндокринной ткани поджелудочной железы структурно организована в виде внешнего эпителия. Наряду с внешним эпителием, встречаются свободно расположенные панкреатические островки и скопления эндокринных клеток, связанные с протоками. Клеточный состав эндокринных клеток представлен А-, В- и D-клетками.
Поджелудочная железа двоякодышащей рыбы протоптеруса в отличие от кистеперых рыб имеет эндокринную часть поджелудочной железы, представленную не внешним эпителием, а панкреатическими островками, окруженными соединительнотканной капсулой. Клеточный состав их: А-, В- и негранулированные клетки.
Надотряд костистых рыб является боковой ветвью эволюции костных рыб, однако знание морфологии эндокринной части поджелудочной железы его представителей важно для понимания общих закономерностей структурной эволюции эндокринной ткани поджелудочной железы. Новые данные по этому вопросу получены В. В. Ягловым. Выяснено, что эндокринная часть поджелудочной железы костистых рыб организована в форме панкреатических островков. У некоторых видов (например, у горбуши) наряду с панкреатическими островками имеются еще «тельца Брокмана». Тельца Брокмана — уникальная форма структурной организации эндокринной ткани поджелудочной железы позвоночных. Это сравнительно крупные образования, окруженные плотной соединительнотканной капсулой, через которую в них входят кровеносные сосуды и нервы. Клеточный состав эндокринной части поджелудочной железы костистых рыб — А-, В- и D-клетки.
Таким образом, у костистых рыб мы наблюдаем переход к новой эволюционной форме организации эндокринной ткани поджелудочной железы — к панкреатическим островкам.
Земноводные, или амфибии. Земноводные берут свое начало от древних форм кистеперых рыб и являются, по существу, первыми примитивными наземными позвоночными. У всех видов амфибий (хвостатых — например, тритон, бесхвостых — травяная лягушка) эндокринная часть поджелудочной железы структурно организована в виде панкреатических островков, образованных А-, В- и D-клетками. Помимо панкреатических островков, в поджелудочной железе травяной лягушки и серой жабы описаны эндокринные клетки, в составе эпителия протоков, что рассматривается как остатки внешнего эпителия и подтверждает филогенетическое родство амфибий и кистеперых рыб.
Согласно данным В. В. Яглова панкреатические островки амфибий построены по общему плану. В центре островка находится синусоидальный капилляр, вокруг которого располагаются В-клетки. А- и D-клетки лежат на периферии островков. Если секреторные гранулы в В-клетках скапливаются в отделах цитоплазмы, обращенных к капилляру, и выделяют гормон непосредственно в кровь — «прямой» путь транспорта гормона, то А- и D-клетки, как правило, не образуют скоплений секреторных гранул и выделяют гормоны в межклеточные щели по микроапокриновому способу. По межклеточным щелям гормоны поступают в перикапиллярные пространства и далее в капилляры — «непрямой» путь транспорта гормонов. Функциональное значение «непрямого» пути транспорта гормона В. В. Яглов усматривает в возможности внеклеточного депонирования секреторных гранул, после чего гормон может быть легко мобилизован для нужд организма в адекватных количествах. Помимо этого, контакт гормона с рецепторами плазматической мембраны железистых клеток в процессе транспорта гормона по межклеточным щелям может оказывать регулирующее влияние на процессы секретообразования.
Пресмыкающиеся, или рептилии. Все изученные виды класса пресмыкающихся могут быть подразделены на три группы. Первая — самая многочисленная — представлена некоторыми видами отряда чешуйчатых (гадюковые и ужеобразные, змеи, агамы и настоящие ящерицы). У них эндокринная часть поджелудочной железы образована панкреатическими островками, состоящими из А-, В- и D-клеток. В эволюционном плане эти рептилии по строению эндокринной части поджелудочной железы занимают положение, промежуточное между амфибиями и млекопитающими.
Вторая группа представлена отрядом черепах. У черепах эндокринная часть поджелудочной железы — это не только панкреатические островки, но также тяжи А-клеток, которые располагаются вне панкреатических островков и не связаны с панкреатическими протоками. Последнее рассматривается, как прообраз «темных» панкреатических островков птиц, как признак расселения контринсулярного компонента за пределы панкреатического островка.
Третья группа представлена подотрядом ящерицы, семейством варанов, у кеторых имеются панкреатические островки и внешний эпителий, образованный Б-клетками.
Таким образом, животные этой группы сочетают признаки, свойственные рептилиям, а также хрящевым и кистеперым рыбам. Указанные различия, вероятно, являются отражением экологической адаптации и различных путей эволюционного развития рептилий.
Птицы. В поджелудочной железе птиц обнаруживаются эволюционно новые формы структурной организации эндокринной ткани. Эндокринная часть поджелудочной железы птиц представлена панкреатическими островками трех типов: «темными», «светлыми» и «смешанными». «Темные» островки состоят из А- и единичных D-клеток. «Светлые» — из В- и единичных D-клеток, «смешанные» — из А-, В- и D-клеток. В функциональном отношении «темные» островки представляют контринсулярный компонент эндокринной ткани поджелудочной железы птиц. Сам факт существования «темных» и «светлых» панкреатических островков свидетельствует о структурном разъединении инсулярного и контринсулярного компонентов эндокринной части поджелудочной железы. Это разъединение, однако, носит частичный характер, о чем свидетельствует наличие «смешанных» островков, содержащих как инсулярный, так и контринсулярный компоненты. Это является первой особенностью структурной организации панкреатических островков у птиц.
Вторая особенность — низкий показатель соотношения В- и А-клеток в панкреатических островках. Индекс В/А, по данным Ю. К. Елецкого и В. В. Яглова, колеблется у птиц от 0,39 до 0,40, т. е. количественно А-клетки приблизительно в 4 раза преобладают над В-клетками. Согласно биохимическим данным содержание глюкагона (гормона А-клеток) в поджелудочной железе птиц превышает содержание инсулина (гормона В-клеток) приблизительно в 10 раз.
Оценивая факторы, обусловившие такой путь эволюции эндокринной ткани поджелудочной железы птиц, следует выделить особенности метаболизма. Прежде всего это необходимость поддержания высокого уровня сахара в крови, что, в свою очередь, необходимо для интенсивного течения метаболизма и поддержания высокой температуры тела. Во-вторых, при совершении длительных перелетов, особенно в период миграций, требуется оперативная мобилизация глюкозы и жирных кислот из гликогена и триглицеридов жировых депо, что осуществляется под влиянием глюкагона. Возможно, что структурное разъединение инсулярного (обеспечивающего депонирование глюкозы в форме гликогена и свободных жирных кислот в форме триглицеридов) и контринсулярного, оказывающего противоположное действие, компонентов обеспечивает возможность более целенаправленного регулирования энергетического обмена в зависимости от потребностей организма.
Ю, К. Елецкий и В. В. Яглов отмечают еще одну важную особенность структурной организации «темных» островков птиц. Это наличие в них локальных расширений межклеточных щелей и относительная бедность кровеносными сосудами. Авторы считают, что цитоплазматические выросты поверхности экзокринных панкреацитов, окружающих «темные» островки, перекрывая межклеточные щели, обеспечивают внеклеточное депонирование клюкагона и адекватное выведение его в сосудистое русло при возникающих потребностях.
Млекопитающие. У всех изученных представителей отрядов зайцеобразных, хищных, грызунов, рукокрылых, непарно- и парнокопытных, китообразных и приматов эндокринная часть поджелудочной железы организована в форме панкреатических островков, хотя у многих представителей отмечена и внеостровковая локализация отдельных эндокринных клеток (например, в составе эпителия протоков, в составе концевых отделов экзокринной части). В эволюционном отношении панкреатические островки млекопитающих являются «смешанными» островками, состоящими из А-, В-иD-клеток. В количественном отношении преобладающим клеточным типом являются В-клетки. Здесь, вероятно, следует указать на существование экологической детерминации в отношении преобладания того или иного клеточного типа в панкреатическом островке. Так, к примеру, у дельфина, местом обитания которого является вода, панкреатические островки по содержанию А- и В-клеток напоминают островки костистых рыб, у которых индекс В/А имеет значения от 0,66 до 1,85.
Подводя итог нашего краткого обзора основных этапов эволюции эндокринной части поджелудочной железы, можно заключить, что у бесчерепных все клеточные элементы, гомологичные клеткам поджелудочной железы, локализуются в эпителии слизистой оболочки кишечника и его печеночном выросте. У круглоротых за счет миграции части железистых клеток формируется инсулярный аппарат (скопления эндокринных клеток, фолликулы Лангерганса) в собственном слое слизистой или среди соединительной ткани, окружающей кишечник. Клетки же, гомологичные клеткам экзокринной части поджелудочной железы, локализуются в слизистой кишечника. У хрящевых рыб благодаря выселению остальных гомологов формируется поджелудочная железа, состоящая из экзо- и эндокринной частей. Первой формой организации эндокринной части железы является внешний эпителий. В ходе дальнейшей эволюции (амфибии, рептилии, птицы, млекопитающие) стабилизировалась новая форма структурной организации эндокринной части поджелудочной железы — панкреатические островки (рис. 13).

Схема этапов эволюции структурной организации эндокринной части поджелудочной железы позвоночных…
Источник
Эмбриональное развитие поджелудочной железы
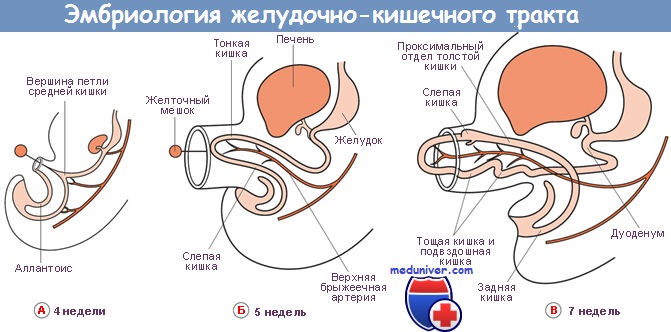
Поджелудочная железа развивается из отдельных зачатков — вентрального и дорсального, — образующихся из соединения эмбриональных передней кишки со средней. Дорсальный зачаток увеличивается влево и формирует основную массу зрелой железы.
Вентральный зачаток, тесно связанный с развивающимся общим желчным протоком, первоначально отходит от двенадцатиперстной кишки. Он соединяется с дорсальной системой только на седьмой неделе внутриутробного развития, после ротации позади эмбриональной двенадцатиперстной кишки, и расположен каудально от дорсального зачатка.
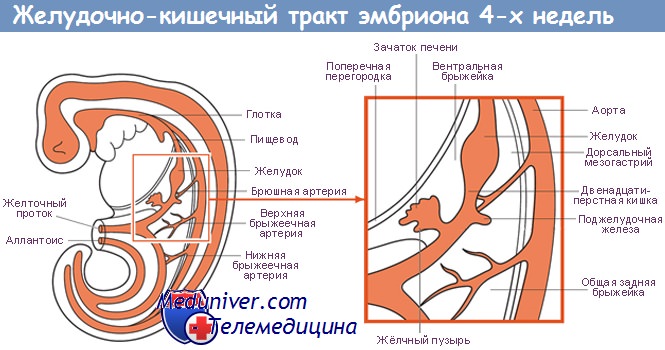
Обе части первичной поджелудочной железы содержат аксиальные протоки: дорсальный проток развивается из стенки двенадцатиперстной кишки, а вентральный — из общего желчного протока. При их слиянии вентральный проток (вирзунгов) продолжается дорсальным протоком (санториниев), образуя главный панкреатический проток.
Таким образом, общий желчный и панкреатический протоки открываются в двенадцатиперстную кишку на большом сосочке, тогда как часть дорсального протока внутри головки поджелудочной железы становится более или менее рудиментарной и открывается в двенадцатиперстную кишку проксимально от большого сосочка на небольшом добавочном или малом сосочке.
Полное отсутствие слияния систем двух протоков приводит к развитию разделенной поджелудочной железы. Такая аномалия встречается приблизительно у 5% людей и может быть предрасполагающим фактором панкреатита. Также возможно неполное слияние, при котором санториниев проток остается главным.
Недостаточный поворот тела вентрального зачатка по оси с устьем его протока может привести к развитию кольцевидной поджелудочной железы, окружающей вторую часть двенадцатиперстной кишки, что может вызвать обструкцию двенадцатиперстной кишки.
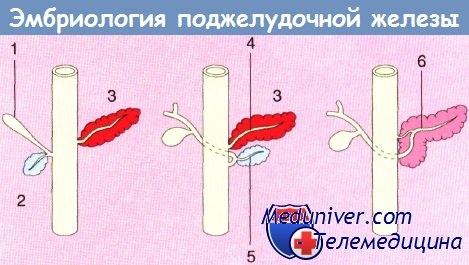
Схема, показывающая анатомию развивающейся поджелудочной железы:
1 — билиарный зачаток; 2 — вентральный;
3 — дорсальный; 4 — санториниев проток;
5 — вирзунгов проток; 6 — главный панкреатический проток.
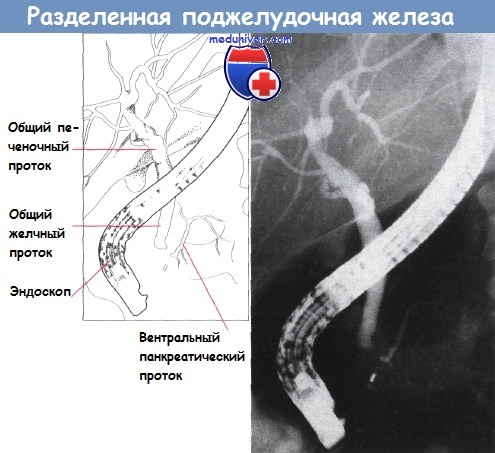
Разделенная поджелудочная железа. Определяются протоки в вентральной части железы.
Видно их отсутствие справа от позвоночника.
Хорошо заметно близкое расположение к поджелудочной железе общего желчного протока.
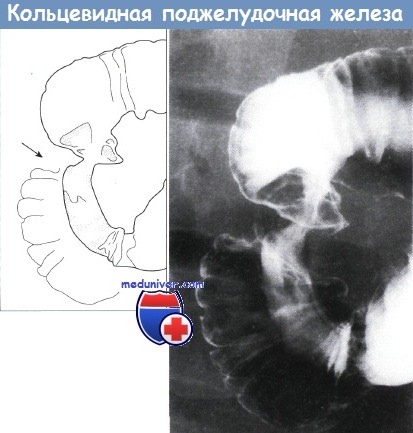
Верхний отдел желудочно-кишечного тракта, кольцевидная поджелудочная железа.
Наблюдается очаговое циркулярное ровное сужение середины второй части двенадцатиперстной кишки.
Дуоденальные складки и слизистая оболочка в остальном нормальные. Рентгенограмма с двойным контрастированием.
— Также рекомендуем «Острый панкреатит — диагностика»
Оглавление темы «Заболевания желудочно-кишечного тракта»:
- Сифилис заднего прохода — диагностика
- Гонорея прямой кишки — диагностика
- Предраковые состояния анальной области — диагностика
- Злокачественные опухоли анального канала — диагностика
- Врожденные аноректальные аномалии — диагностика
- Поджелудочная железа в норме — анатомия, методы обследования
- Эмбриональное развитие поджелудочной железы
- Острый панкреатит — диагностика
- Хронический панкреатит — диагностика
- Наследственный панкреатит — диагностика
Источник
