
Назовите пептидазы поджелудочной железы
За сутки образуется 1,5-2,5 л панкреатического сока, рН — 7,5-8,8. Высокое содержание ионов бикарбоната — обеспечивают нейтрализацию кислого желудочного содержимого.
Специфические вещества поджелудочного сока:
1. Панкреатический калликреин (активирует моторику, расширяет сосуды).
2. Ингибитор трипсина (блокирует активацию трипсина внутри железы).
Панкреатический сок содержитвсе группы ферментов.
Протеазыподжелудочного сока(эндо- и экзопептидазы):
а). Эндопептидазы расщепляют внутренние пептидные связи.
1. Трипсин — расщепляет связи между аргинином и лизином, активируется энтерокиназой и аутокаталитически.
2. Химотрипсин — расщепляет связи тирозина, триптофана, фенилаланина , активируется трипсином.
3. Панкреопептидаза Е (эластаза) — расщепляет эластические белки.
б ). Экзопептидазы (карбокси- и аминопептидазы) расщепляют конечные связи, освобождая аминокислоты с «С»-конца пептида (СООН) и с » N «-конца пептида (NH3).
Липазыподжелудочного сока:
Липаза поджелудочной железы вырабатывается в активном состоянии, действует на эмульгированные желчью жиры, расщепляя их до глицерина и ВЖК.
Фосфолипаза А вырабатывается в неактивном состоянии, активируется трипсином,расщепляет фосфолипиды до жирных кислот.
Карбогидразыподжелудочного сока: альфа-амилаза, альфа-глюкозидаза.
Нуклеазы(класс фосфодиэстераз):рибонуклеаза, дезоксинуклеаза.
Желчь :Объем суточной секреции — 0,5-1 л., рН — 7,8-8,6.
1. Желчь не содержит ферментов.
2. Специфические вещества: желчные кислоты и желчные пигменты:
— билирубин — основной пигмент у человека, придает коричневую окраску;
— биливердин — в основном в желчи травоядных животных (зеленый цвет).
Желчные кислоты,играют в пищеварении ведущую роль:
— эмульгируют жиры,
— усиливают активность поджелудочной липазы,
— обеспечивают всасывание нерастворимых в воде веществ (ЖК, холестерин, витамины (А, D , Е, К) и соли Са+2),
— способствуют ресинтезу триглицеридов в энтероцитах.
Сок тонкой кишки — 2,5 л. в сутки, рН — 7,2-7,5.
Специфическими веществами кишечного сока являются щелочные продукты.
Ферменты : всего – более 20 ферментов. Наиболее значимые:
Протеазы кишечного сока:
1. Энтерокиназа — фермент, активирующий трипсиноген.
2. Три- и дипептидазы(эрипсины)- расщепляют пептиды на аминокислоты.
Липазысодержатся в незначительном количестве (липаза, фосфолипаза).
Карбогидразыкишечного сока: альфа-глюкозидаза, бета-галактозидаза ( расщепляет молочный сахар до глюкозы и галактозы), сахараза, лактаза, гамма-амилаза.
Сок толстой кишки —рН сока — 8,5-9,0.
Специфические вещества – слизь, обеспечивает формирование каловых масс.
Собственных ферментов сок толстой кишки не содержит.
Состав сока толстой кишки во многом зависит от его микрофлоры,которая:
1. Предохраняет организм хозяина от внедрения и размножения патогенных микроорганизмов (предотвращает процессы гниения (белки) и брожения (углеводы)).
2. Участвует в разложении компонентов пищеварительных секретов (ферментов, желчных кислот иклетчатки).
3. Способна синтезировать витамин К и некоторые витамины группы В.
Дата добавления: 2015-03-27; просмотров: 975; Опубликованный материал нарушает авторские права? | Защита персональных данных | ЗАКАЗАТЬ РАБОТУ
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Учись учиться, не учась! 10322 — | 7850 — или читать все…
Читайте также:
Источник
В
пищевых продуктах содержание свободных
аминокислот очень мало. Подавляющее их
количество входит в состав белков,
которые гидролизуются в ЖКТ под действием
ферментов протеаз (пептидщцролаз).
Субстратная специфичность этих ферментов
заключается в том, что каждый из них с
наибольшей скоростью расщепляет
пептидные связи, образованные определёнными
аминокислотами. Протеазы, гидролизующие
пептидные связи внутри белковой молекулы,
относят к группе эндопептидаз. Ферменты,
относящиеся к группе экзопептидаз,
гидролизуют пептидную связь, образованную
концевыми аминокислотами. Под действием
всех протеаз ЖКТ белки пищи распадаются
на отдельные аминокислоты, которые
затем поступают в клетки тканей.
Переваривание
белков в желудке
Желудочный
сок — продукт нескольких типов клеток.
Обкладочные (париетальные) клетки стенок
желудка образуют соляную кислоту,
главные клетки секретируют пепсиноген.
Добавочные и другие клетки эпителия
желудка выделяют муцинсодержащую слизь.
Париетальные клетки секретируют в
полость желудка также гликопротеин,
который называют «внутренним фактором»
(фактором Касла). Этот белок связывает
«внешний фактор» — витамин В12,
предотвращает его разрушение и
способствует всасыванию.
Образование
и роль соляной кислоты.
Основная
пищеварительная функция желудка
заключается в том, что в нём начинается
переваривание белка. Существенную роль
в этом процессе играет соляная кислота.
Белки, поступающие в желудок, стимулируют
выделение гистамина и
группы белковых гормонов —гастринов которые,
в свою очередь, вызывают секрецию НСI и
профермента — пепсиногена. Источником
Н+ является
Н2СО3,
которая образуется в обкладочных клетках
желудка из СО2,
диффундирующего из крови, и Н2О
под действием фермента карбоангидразы
(карбонатдегидра-тазы):
Н2О
+ СО2 →
Н2СО3 →
НСО3— +
H+
Диссоциация
Н2СО3 приводит
к образованию бикарбоната, который с
участием специальных белков выделяется
в плазму в обмен на С1-,
и ионов Н+,
которые поступают в просвет желудка
путём активного транспорта, катализируемого
мембранной Н+/К+-АТФ-азой.
При этом концентрация протонов в просвете
желудка увеличивается в 106 раз.
Ионы Сl- поступают
в просвет желудка через хлоридный канал.
Концентрация НСl в желудочном соке может
достигать 0,16 М, за счёт чего значение
рН снижается до 1,0-2,0. Приём белковой
пищи часто сопровождается выделением
щелочной мочи за счёт секреции большого
количества бикарбоната в процессе
образования НСl. Под действием НСl
происходит денатурация белков пищи, не
подвергшихся термической обработке,
что увеличивает доступность пептидных
связей для протеаз. НСl обладает
бактерицидным действием и препятствует
попаданию патогенных бактерий в кишечник.
Кроме того, соляная кислота активирует
пепсиноген и создаёт оптимум рН для
действия пепсина.
Механизм
активации пепсина.
Под
действием гастринов в главных клетках
желудочных желёз стимулируются синтез
и секреция пепсиногена — неактивной
формы пепсина. Пепсиноген — белок,
состоящий из одной полипептидной цепи
с молекулярной массой 40 кД. Под действием
НСl он превращается в активный пепсин
(молекулярная масса 32,7 кД) с оптимумом
рН 1,0-2,5. В процессе активации в результате
частичного протеолиза от N-конца молекулы
пепсиногена отщепляются 42 аминокислотных
остатка, которые содержат почти все
положительно заряженные аминокислоты,
имеющиеся в пепсиногене. Таким образом,
в активном пепсине преобладающими
оказываются отрицательно заряженные
аминокислоты, которые участвуют в
конформационных перестройках молекулы
и формировании активного центра.
Образовавшиеся под действием НСl активные
молекулы пепсина быстро активируют
остальные молекулы пепсиногена
(аутокатализ). Пепсин в первую очередь
гидролизует пептидные связи в белках,
образованные ароматическими аминокислотами
(фенилаланин, триптофан, тирозин) и
несколько медленнее — образованные
лейцином и дикарбоновыми аминокислотами.
Пепсин — эндопептидаза, поэтому в
результате его действия в желудке
образуются более короткие пептиды, но
не свободные аминокислоты.
Переваривание
белков в кишечнике.
Желудочное
содержимое (химус) в процессе переваривания
поступает в двенадцатиперстную кишку.
Низкое значение рН химуса вызывает в
кишечнике выделение белкового гормона
секретина, поступающего в кровь. Этот
гормон в свою очередь стимулирует
выделение из поджелудочной железы в
тонкий кишечник панкреатического сока,
содержащего НСО3-,
что приводит к нейтрализации НСl
желудочного сока и ингибированию
пепсина. В результате рН резко возрастает
от 1,5-2,0 до ∼7,0.
Поступление пептидов в тонкий кишечник
вызывает секрецию другого белкового
гормона — холецистокинина, который
стимулирует выделение панкреатических
ферментов с оптимумом рН 7,5-8,0. Под
действием ферментов поджелудочной
железы и клеток кишечника завершается
переваривание белков.
Активация
панкреатических ферментовВ
поджелудочной железе синтезируются
проферменты ряда протеаз: трипсиноген,
химотрипсиноген, проэластаза,
прокарбоксипептидазы А и В. В кишечнике
они путём частичного протеолиза
превращаются в активные ферменты
трипсин, химотрипсин, эластазу и
карбоксипептидазы А и В.
Активация
трипсиногена происходит
под действием фермента эпителия кишечника
энтеропептидазы. Этот фермент отщепляет
с N-конца молекулы трипсиногена гексапептид
Вал-(Асп)4-Лиз.
Изменение конформации оставшейся части
полипептидной цепи приводит к формированию
активного центра, и образуется активный
трипсин. Последовательность Вал-(Асп)4-Лиз
присуща большинству известных
трипсиноге-нов разных организмов — от
рыб до человека.
Образовавшийся
трипсин активирует
химотрипсиноген, из
которого получается несколько активных
ферментов (рис. 9-3). Химотрипсиноген
состоит из одной полипептидной цепи,
содержащей 245 аминокислотных остатков
и пяти дисульфидных мостиков. Под
действием трипсина расщепляется
пептидная связь между 15-й и 16-й
аминокислотами, в результате чего
образуется активный π-химотрипсин.
Затем под действием π-химотрипсина
отщепляется дипептид сер(14)-арг(15), что
приводит к образованию δ-химотрипсина.
Отщепление дипептида тре(147)-арг(148)
завершает образование стабильной формы
активного фермента — α-химотрипсина,
который состоит из трёх полипептидных
цепей, соединённых дисульфидными
мостиками. Остальные проферменты
панкреатических протеаз (проэластаза
и прокарбоксипептидазы А и В) также
активируются трипсином путём частичного
протеолиза. В результате образуются
активные ферменты — эластаза и
карбокси-пептидазы А и В.
Специфичность
действия протеаз.
Трипсин
преимущественно гидролизует пептидные
связи, образованные карбоксильными
группами аргинина и лизина. Химотрипсины
наиболее активны в отношении пептидных
связей, образованных карбоксильными
группами ароматических аминокислот
(Фен, Тир, Три). Карбоксипептидазы А и В
— цинксодержащие ферменты, отщепляют
С-концевые остатки аминокислот. Причём
карбоксипептидаза А отщепляет
преимущественно аминокислоты, содержащие
ароматические или гидрофобные радикалы,
а карбоксипептидаза В — остатки аргинина
и лизина. Последний этап переваривания
— гидролиз небольших пептидов, происходит
под действием ферментов аминопептидаз
и дипептидаз, которые синтезируются
клетками тонкого кишечника в активной
форме.
Аминопептидазы последовательно
отщепляют N-концевые аминокислоты
пептидной цепи. Наиболее известна
лейцинаминопептидаза — Zn2+-
или Мn2+-содержащий
фермент, несмотря на название, обладающий
широкой специфичностью по отношению
к N-концевым аминокислотам.Дипептидазы расщепляют
дипептиды на аминокислоты, но не
действуют на трипептиды.
В результате последовательного действия
всех пищеварительных протеаз большинство
пищевых белков расщепляется до свободных
аминокислот.
Экзопептидазы(экзопротеиназы)
—ферменты,
гидролизующие белки, отщепляяаминокислотыот
концапептида:карбоксипептидазы—
от C-конца,аминопептидазы—
от N-конца,дипептидазырасщепляют
дипептиды. Экзопептидазы синтезируются
в клеткахтонкого
кишечника(аминопептидазы,
дипептидазы) и вподжелудочной
железе(карбоксипептидаза).
Функционируют эти ферменты внутриклеточно
в кишечномэпителиии,
в небольшом количестве, в просветекишечника.
Эндопептидазы(эндопротеиназы) —протеолитические
ферменты(пепсин,трипсин,химотрипсин),
расщепляющиепептидные
связивнутрипептидной
цепи. С наибольшей скоростью
ими гидролизуются связи, образованные
определённымиаминокислотами.
Эндопептидазы синтезируются в
видепроферментов,
активируемых затем при помощи
избирательногопротеолиза.
Таким образом клетки, секретирующие
эти ферменты защищают собственные белки
от разрушения. От действия ферментовклеточную
мембрануклеток животных
защищает также поверхностный
слойолигосахаридов—гликокаликс,
а вкишечникеижелудке—
слой слизи.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
единица | 9.1. Роль 9.2. Переваривание 9.3. Трансаминирование |
Модульная | 9.4. Обмен 9.5. Орнитиновый 9.6. Гипераммониемия 9.7. Пути 9.8. Биосинтез |
Модульная | 9.9. Обмен 9.10. Обмен 9.11. Обмен 9.12. Заболевания, 9.13. Биогенные |
ЕМА 9.1. РОЛЬ
БЕЛКОВ В ПИТАНИИ. АЗОТИСТЫЙ БАЛАНС
В организме
человека содержится примерно 15 кг
белков. Количество свободных аминокислот
составляет около 35 г. Ежесуточно в
организме распадается до аминокислот
почти 400 г белков и столько же синтезируется.
1. Основным
источником аминокислот для человека
являются пищевые белки. Суточная норма
потребления белков составляет в среднем
около 100 г.
20 α-аминокислот,
которые встречаются в белках организма,
можно разделить на четыре группы:
• заменимые
аминокислоты — Ала, Асп, Асн, Глу, Глн,
Про, Гли, Сер —
синтезируются в необходимых количествах
в организме;
• незаменимые
аминокислоты — Вал, Лей, Иле, Мет, Фен,
Три, Лиз, Тре —
не могут синтезироваться в организме
и должны поступать с пищей;
• частично
заменимые аминокислоты — Гис, Арг —
синтезируются очень медленно, в
количествах, не покрывающих потребности
организма, особенно в детском возрасте;
• условно
заменимые аминокислоты — Цис, Тир —
синтезируются из незаменимых аминокислот
Мет и Фен соответственно.
2. Полноценность
белкового питания зависит
от аминокислотного состава белков и
определяется наличием незаменимых
аминокислот.Отсутствие
в пищевых белках незаменимых аминокислот
(даже одной) нарушает их синтез в
организме. Обновление белков в различных
тканях происходит с разной скоростью.
Так, белок соединительной ткани коллаген
обновляется полностью за 300 дней, а
белки системы свертывания крови — от
нескольких минут до нескольких дней.
3. Большая
часть свободных аминокислот используется
для синтеза собственных белков организма.
Кроме того, из аминокислот синтезируется
большое количество биологически
активных молекул:
• биогенные
амины (медиаторы); некоторые аминокислоты
сами являются нейромедиаторами —
например, глицин и глутамат;
• гормоны
белковой природы;
• гем,
креатин, карнитин и другие азотсодержащие
соединения.
Аминокислоты
подвергаются реакции дезаминирования;
образовавшиеся безазотистые остатки
используются для синтеза глюкозы,
кетоновых тел или окисляются до СО2 и
Н2О.
Азот аминокислот
выводится из организма почками в виде
мочевины или аммонийных солей.
Аминокислоты и белки содержат до 95%
всего азота организма.
4. Азотистый
баланс —
разница между количеством азота,
поступающего с пищей, и количеством
азота, выделяемого почками в виде
мочевины и азотистых солей. Он является
показателем состояния белкового и
аминокислотного обмена.
Азотистый
баланс может
быть:
• положительным —
у детей, беременных женщин, при увеличении
мышечной массы у спортсменов и больных,
выздоравливающих после тяжелой болезни,
что свидетельствует о преобладании
синтеза белков и роста тканей над их
распадом;
• отрицательным —
при тяжелых заболеваниях, голодании,
старении, что свидетельствует об
усилении процессов распада белков;
• равным
нулю (азотистое равновесие) — у
здоровых взрослых людей при нормальном
питании.
1. При
переваривании происходит гидролиз
пищевых белков до свободных аминокислот.
Процесс переваривания начинается в
желудке и продолжается в тонком кишечнике
под действием ферментов пептидгидролаз
(пептидаз). Основные
пептидазы синтезируются в клетках
желудка, поджелудочной железы и кишечника
(рис. 9.1). В желудке белки
пищи денатурируются и гидролизуются
с образованием олигопептидов.
Вкишечнике панкреатические
пептидазы продолжают гидролиз
олигопептидов до ди- и трипептидов и
свободных аминокислот. Короткие пептиды
расщепляются до свободных аминокислот
в пристеночном
слое или
в клетках кишечного эпителия. Затем
происходит их всасывание.
Все пептидазы, в
зависимости от места расположения в
пептиде гидролизуемой связи, делятся
на
эндопептидазы и экзопептидазы:
• эндопептидазы —
расщепляют пептидные связи, удаленные
от концов пептидной цепи (пепсин,
трипсин, химотрипсин, эластаза);
• экзопептидазы
— гидролизуют
пептидные связи, образованные N- и
С-концевыми аминокислотами (аминопептидаза,
карбоксипептидазы А и В),
а также расщепляют ди- и трипептиды.
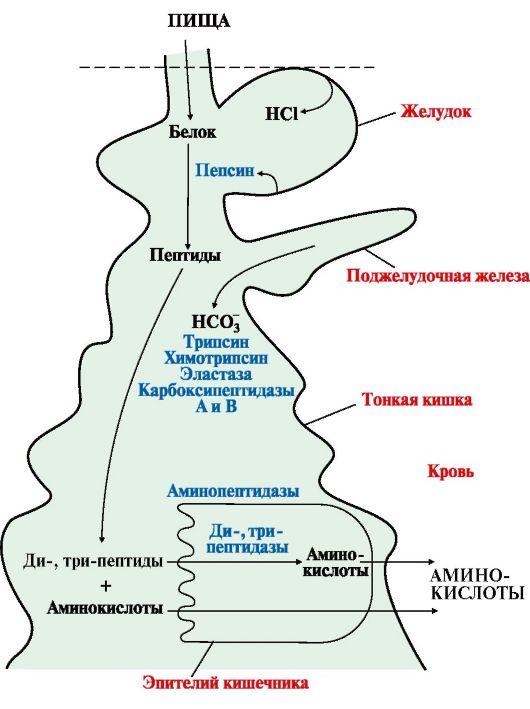 Рис.
Рис.
9.1. Переваривание белков в желудочно-кишечном
тракте
Ферменты,
участвующие в переваривании белков,
обладают относительной субстратной
специфичностью и гидролизуют пептидные
связи; при этом каждая пептидаза
преимущественно расщепляет связи,
образованные определенными аминокислотами
(табл. 9.1).
2. Желудочные
и панкреатические пептидазы вырабатываются
в неактивной форме (проферменты), секретируются
к месту действия, где активируются путем
частичного протеолиза (отщепление
пептида различной длины, чаще с N-конца
молекулы профермента). Место
синтеза проферментов (клетки
слизистой оболочки желудка, поджелудочная
железа) и место
их активации (полость
желудка, тонкого кишечника)пространственно
разделены. Такой
механизм образования активных ферментов
необходим для защиты секреторных клеток
желудка и поджелудочной железы от
самопереваривания.
Таблица
9.1. Характеристика протеолитических
ферментов желудочно-кишечного тракта
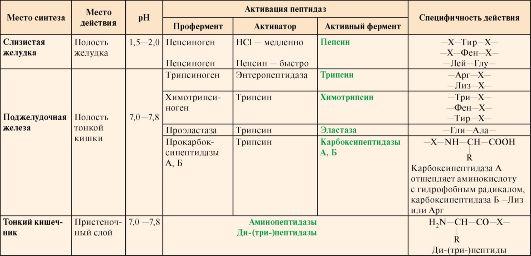 Примечание: X
Примечание: X
— любая аминокислота
Преждевременная
активация проферментов в секреторных
клетках происходит при:
• язвенной
болезни желудка —
пепсиноген превращается в пепсин в
клетках слизистой желудка;
• остром
панкреатите —
трипсиноген превращается в трипсин в
клетках поджелудочной железы и активирует
остальные панкреатические пептидазы.
3. Слизистая
оболочка желудка вырабатывает следующие
факторы, необходимые для переваривания
белков:
• пепсиноген —
синтезируется в главных клетках;
• соляную
кислоту —
вырабатывается обкладочными клетками.
Гидролиз белков в
желудке происходит
под действием пепсина.
Профермент
пепсиноген при поступлении пищи
секретируется в полость желудка, где
в два этапа происходит его активация:
• с помощью
НС! — медленно;
• аутокаталитически
— быстро, уже имеющимся пепсином. Соляная
кислота желудочного
сока выполняет следующие функции:
• .участвует
в активации пепсиногена;
• создает
оптимум рН для пепсина;
• оказывает
бактерицидное действие;
• денатурирует
белки пищи.
Значение рН
желудочного сока в норме составляет
1,5-2,0. Определение
кислотности желудочного сока используется
для диагностики различных заболеваний
желудка (табл. 9.2).
Таблица
9.2. Компоненты желудочного сока в норме
и при патологических состояниях
• 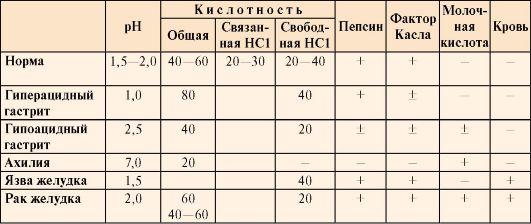 Общая
Общая
кислотность желудочного сока —
это совокупность всех кислотореагирующих
веществ желудочного сока, представляющая
собой секрет желудка, собираемый в
течение 1 часа (предварительно отбирают
секрет, ранее содержащийся в желудке).
• Связанная
соляная кислота —
это соляная кислота, связанная с белками
и продуктами их переваривания.
• Свободная
соляная кислота —
это соляная кислота, не связанная с
другими молекулами.
• Общая
кислотность желудочного сока выражается
в титрационных единицах (ТЕ) и
измеряется количеством 0,1 М NaOH в
миллилитрах, затраченным на титрование
100 мл желудочного сока в присутствии
определенных индикаторов.
• Кислотность
желудочного сока в
норме составляет:
общая 40-60 ТЕ;
связанная
HCl 20-30 ТЕ;
свободная
HCl 20-40 ТЕ.
При диагностике
заболеваний желудка помимо биохимических
анализов обязательно проводят
рентгенологические и эндоскопические
исследования, иногда биопсию.
В слизистой
оболочке желудка вырабатывается
также внутренний
фактор (фактор Касла), который
представляет собой белок, способствующий
всасыванию витамина В12 в
тонкой кишке. Отсутствие этого витамина
часто приводит к развитию анемии.
Молочная
кислота в
норме в желудочном соке отсутствует.
Образуется при уменьшении содержания
или отсутствии свободной соляной
кислоты в результате усиленного
размножения молочнокислых бактерий
или при злокачественных опухолях
желудка.
НС1 и пепсин
способны разрушать клетки эпителия
желудка. В норме это не происходит
благодаря наличию защитных факторов
слизистой оболочки, таких, как:
• образование
на поверхности слизи, содержащей
гетерополисахариды, которые не являются
субстратами пептидгидролаз;
• секреция
эпителиальными клетками ионов НСО3-,
создающих в пристеночном слое менее
агрессивную среду с рН 5,0-6,0, в которой
пепсин неактивен. Кроме того, клетки
поврежденного эпителия обладают
способностью к быстрой регенерации.
Пепсин
гидролизует пептидные
связи, образованные аминогруппой или
карбоксильной группой ароматических
аминокислот (см. табл. 9.1):
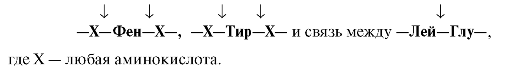 4.
4.
Переваривание белков в кишечнике происходит
под действием:
• ферментов
поджелудочной железы-трипсина,
химотрипсина, эластазы, карбоксипептидаз
А и Б;
• ферментов
эпителия тонкой кишки — аминопептидазы,
дипептидаз, трипептидаз.
Активная
форма трипсина образуется
в кишечнике при участии фермента
энтеропептидазы, выделяемого клетками
кишечника. Энтеропептидазаотщепляет
от N-конца трипсиногена гексапептид
(рис. 9.2), что приводит к изменению
конформации молекулы и формированию
активного центра трипсина.
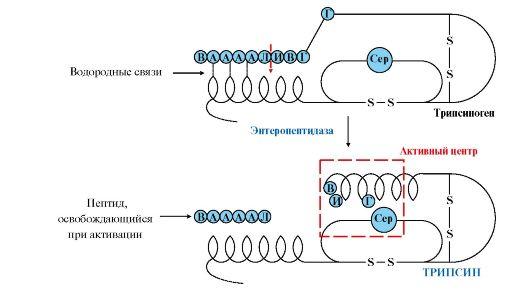 Рис.
Рис.
9.2. Механизм активации трипсиногена.
Пунктирная
стрелка — место гидролиза. Буквами
обозначены аминокислоты (А — асп, Г —
глу, В — вал, Л — лиз, И — иле)
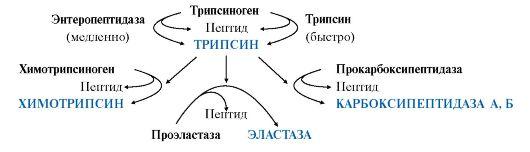
Остальные проферменты
панкреатического сока (химотрипсиноген,
прокарбоксипептидазы А и В, проэластаза)
активируются трипсином.Активация
панкреатических пептидаз в кишечнике
происходит в виде каскада реакций.
 Ферменты
Ферменты
эпителия тонкой кишки синтезируются
в энтероцитах сразу в
активной форме.
• Ферменты,
участвующие в переваривании белков в
кишечнике, обладают
субстратной специфичностью к
определенным аминокислотам в
белке: Трипсин гидролизует
преимущественно пептидные связи,
образованные карбоксильными группами
катионогенных аминокислот:
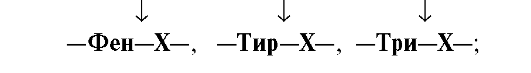
• 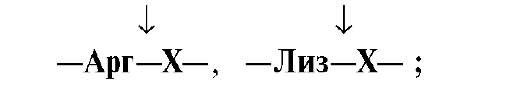 Химотрипсин —
Химотрипсин —
пептидные связи, образованные
карбоксильными группами ароматических
аминокислот:
•  Эластаза —
Эластаза —
связь между -Гли-Ала-
• Карбоксипептидазы отщепляют
С-концевые аминокислоты:
карбоксипептидаза А —
гидрофобные аминокислоты, карбоксипептидаза В
— Лиз и Арг;
• Аминопептидаза —
отщепляет N-концевые аминокислоты;
• Дипептидаза —
гидролизует дипептиды;
• Трипептидаза —
расщепляет трипептиды на отдельные
аминокислоты.
5. Конечным
результатом переваривания белков
является образование свободных
аминокислот, поступающих в клетки
слизистой оболочки кишечника,
путем вторично-активного
транспорта за
счет градиента концентрации натрия
(симпорт). Всасывание L-аминокислот
является активным, требующим затраты
энергии процессом. Транспорт их
осуществляется двумя путями:
• через
воротную систему печени;
• по
лимфатическим сосудам, сообщающимся
с кровью через грудной лимфатический
проток.
Аминокислоты
конкурируют друг с другом за специфические
участки связывания белков-переносчиков.
Так, всасывание лейцина в больших
количествах уменьшает всасывание
изолейцина и валина. В крови максимальная
концентрация аминокислот достигается
через 30-50 минут после приема белковой
пищи. Свободные аминокислоты, в отличие
от белков пищи, лишены видовой
специфичности и не обладают антигенными
свойствами.
Скорость
проникновения аминокислот через
мембраны клеток различается, что
указывает на существование транспортных
систем, обеспечивающих перенос
аминокислот через мембраны. Известно
пять специфических транспортных систем
для переноса определенной группы
близких по строению аминокислот:
• нейтральных
с короткой боковой цепью (аланин,
серин, треонин);
• нейтральных
с длинной или разветвленной боковой
цепью (валин,
лейцин, изолейцин);
• с
катионными радикалами (лизин,
аргинин);
• с
анионными радикалами (глутаминовая
и аспарагиновая кислоты);
• иминокислот (пролин,
оксипролин).
Переносчики
аминокислот первой и пятой групп, а
также переносчик метионина относятся
к числу Na+-зависимых. Независимые от
Na+ переносчики специфичны для некоторых
нейтральных аминокислот (фенилаланин,
лейцин) и аминокислот с катионными
радикалами (лизин).
Одной из
основных транспортных систем для
нейтральных аминокислот
является γ-глутамильный
цикл, который
функционирует в почках, поджелудочной
железе, печени и селезенке, в мозге и
других тканях он содержится в очень
небольших количествах (рис. 9.3).
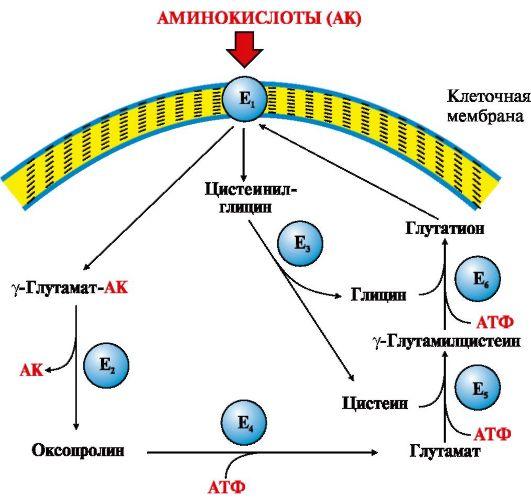 Рис.
Рис.
9.3. γ-Глутамильный цикл:
Е1 —
γ-глутамилтранспептидаза (γ-ГТ); Е2 —
γ-глутамилциклотрансфераза; Е3 —
пептидаза; Е4 —
оксопролиназа; Е5 —
γ-глутамилцистеинсинтетаза; Е6 —
глутатионсинтетаза.
Система
состоит из одного мембранного и пяти
цитоплазматических ферментов. Перенос
аминокислоты внутрь клетки осуществляется
в комплексе с глутамильным остатком
глутатиона под действием фермента
γ-ГТ. Затем аминокислота освобождается,
а γ-глутамильный остаток в несколько
стадий превращается в глутатион, который
способен присоединять следующую
молекулу аминокислоты
Ключевую
роль в работе системы играет мембранный
фермент γ-глутамилтранспептидаза
(γ-ГТ). Этот
гликопротеин катализирует перенос
γ-глутамильной группы с
глутатиона (γ-глутамилцистеинилглицин)
или другого γ-глутамильного пептида
на транспортируемую аминокислоту и
последующий перенос комплекса в клетку.
Глутатион содержится во всех тканях
животных. Для транспорта в клетку одной
молекулы аминокислоты с участием
γ-глутамильного цикла затрачивается
три молекулы АТР.
γ-ГТ в больших
количествах содержится в почках,
поджелудочной железе, печени и других
тканях. Активность фермента в сыворотке
крови в норме составляет 30-50 МЕ/л
(мкмоль/мин-мг) для мужчин и 25-35 МЕ/л для
женщин. Определение активности γ-ГТ в
сыворотке крови используется для
диагностики заболеваний печени и
сердца. γ-ГТ-тест используется также в
качестве маркера рака поджелудочной
железы, печени, предстательной железы
и для обнаружения людей из группы
раннего риска алкоголизма, для контроля
за лечением хронического алкоголизма.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник

