
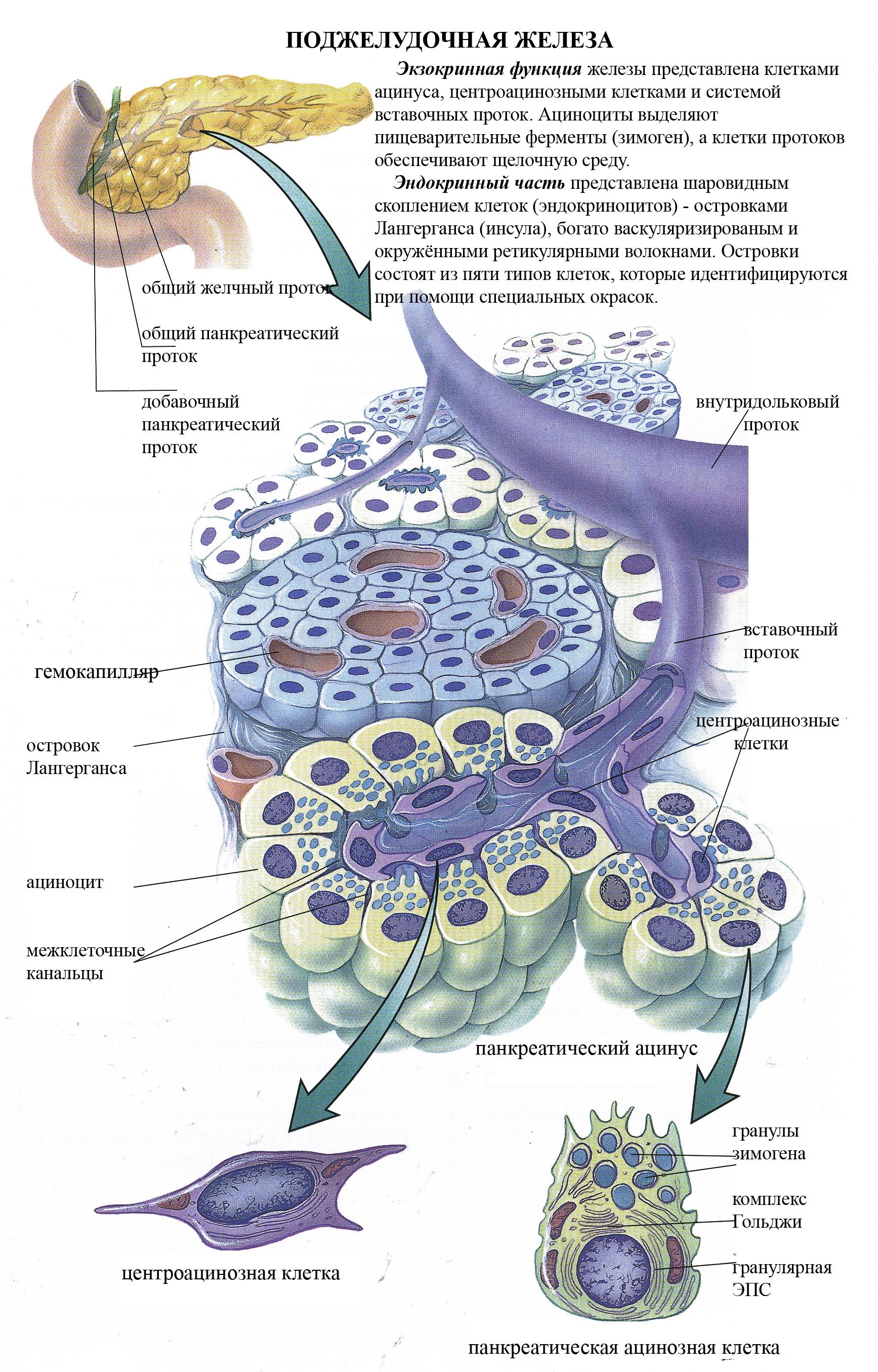
Секреторные включения в клетках поджелудочной железы
К ним относятся белковые, жировые и полисахаридные включения.
Белковые включения. В клетке есть соединения, важность которых определяется тем, что в случае нужды они могут стать предшественниками целого ряда других, жизненно необходимых, для клетки веществ. К таким соединениям относятся аминокислоты. Они могут использоваться в клетке в качестве источников энергии для синтеза углеводов, жиров, гормонов и других метаболитов. Поэтому белковые включения фактически представляют собой своеобразное клеточное сырьё для производства аминокислот.
Судьба белковых включений во всех клетках примерно одинакова. Прежде всего они сливаются с лизосомой, где специальные ферменты расщепляют белки до аминокислот. Последние выходят из лизосом в цитоплазму. Часть из них в цитоплазме взаимодействует с тРНК и в таком виде транспортируется к рибосомам на синтез белка. Другая часть вступает в специальные биохимические циклы, где из них синтезируются жиры, углеводы, гормоны и другие метаболиты. И наконец, аминокислоты принимают участие в энергетическом обмене клетки.
Полисахаридные включения. Для животных клеток и клеток грибов — основным запасным питательным включением является гликоген. Для растений таким включением является крахмал.
Гликоген у человека в основном депонируется в клетках печени и используется не только для нужд самой клетки, но и в качестве энергетических ресурсов для всего организма. В последнем случае гликоген расщепляется в клетке до глюкозы, которая выходит из клетки в кровь и разносится по организму.
Гликоген – это большая ветвистая молекула, состоящая из глюкозных остатков. Специальные внутриклеточные процессы при необходимости отщепляют от молекулы гликогена глюкозные остатки и синтезируют глюкозу. Последняя поступает в кровь и расходуется на нужды клетки. Казалось бы, проще запасти в клетке саму глюкозу, не превращая её в гликоген, тем более что молекула глюкозы растворима и достаточно быстро проходит в клетку через плазматическую мембрану. Однако этому мешает то, что глюкоза также быстро, не задерживаясь, выходит из клетки. Задержать её в клетке в чистом виде практически невозможно. Кроме того, депонирование глюкозы в больших количествах опасно, т.к. это может привести к созданию такого градиента концентраций, что вначале произойдёт набухание клетки из-за поступления воды, а затем и её гибель. Поэтому специальная система ферментов, слегка видоизменив молекулу глюкозы, связывает её с такой же молекулой. Создаётся гигантская ветвистая молекула, состоящая из глюкозных остатков – гликоген. Эта молекула уже нерастворима, как глюкоза, и не способна изменить осмотические свойства клетки.
Жировые включения. Эти включения в гиалоплазме могут находиться в виде капель. Многие растения содержат масла, например подсолнечник, арахис и т.д. Богата жировыми включениями жировая ткань человека, служащая для предохранения организма от потерь тепла, энергетическим депо и как амортизатор при механических воздействиях.
Необходимо отметить, что запасов гликогена в организме среднего взрослого человека достаточно на один день нормальной активности, тогда как запаса жиров хватит на месяц. Если бы главным энергетическим резервом в нашем организме был гликоген, а не жиры, вес тела увеличился бы в среднем на 25 кг.
В некоторых случаях появление жировых включений в клетке является тревожным сигналом неблагополучия. Так, при заболевании дифтерией токсин микроорганизма блокирует утилизацию жирных кислот и они накапливаются в больших количествах в цитоплазме. При этом нарушается метаболизм и клетка гибнет. Чаще всего такие нарушения происходят в клетках сердечной мышцы. Заболевание называется дифтерийный миокардит.
Все питательные включения используются клеткой в моменты интенсивной жизнедеятельности. В эмбриогенезе существует потребность в большом количестве питательных веществ. Поэтому ещё на стадии овогенеза яйцеклетка интенсивно запасает различные питательные вещества (желток и др.) в виде включений, которые обеспечивают прохождение первых этапов эмбрионального развития.
б. Секреторные включения
Различные секреторные гранулы, образующиеся в железистых клетках животных, разнообразны по химической природе и могут быть представлены ионами, ферментами, гормонами, гликопротеинами и т.д., например, пищеварительные ферменты, синтезируемые клетками поджелудочной железы. Сигналом к формированию и опорожнению секреторных включений в поджелудочной железе является приём пищи. До приёма пищи происходит накопление включений в цитоплазме. Определяя количество включений в клетках поджелудочной железы, можно примерно предположить, чьи это клетки – голодного или сытого человека.
в. Пигментные включения
Обеспечивают окраску тканей и органов. В клетках животных часто запасается пигмент меланин. У растений — каротиноиды, флавоноиды.
г. Экскреторные включения
У растений, не имеющих специальных выделительных органов, нередко в вакуолях накапливаются конечные продукты жизнедеятельности в виде солей оксалата или карбоната кальция. В дальнейшем растение сбрасывает листья или кору вместе с такими накопленными кристаллическими включениями.
Источник
Ежедневно поджелудочная железа
секретирует около 1 л литра сока. Сок
поджелудочный железы (ферменты и
бикарбонаты) в ответ на опорожнение
желудка оттекает по длинному выводному
протоку. Этот проток, соединившись с
общим жёлчным протоком, формирует
печёночно-поджелудочную ампулу, которая
открывается на большом дуоденальном
(фатеровом) сосочке в двенадцатиперстную
кишку, будучи окружена жомом из ГМК
(сфинктер Одди). Поступающий в просвет
кишки сок поджелудочной железы содержит
пищеварительные ферменты, необходимые
для переваривания углеводов, белков и
жиров, и большое количество бикарбонатных
ионов, обеспечивающих нейтрализацию
кислого химуса.
Протеолитическиеферменты—
трипсин, химотрипсин, карбоксипептидаза,
эластаза, а также расщепляющие
макромолекулы ДНК и РНК нуклеазы. Трипсин
и химотрипсин расщепляют белки до
пептидов, а карбоксипептидаза расщепляет
пептиды на отдельные аминокислоты.
Протеолитические ферменты находятся
в неактивной форме (трипсиноген,
химотрипсиноген и прокарбоксипептидаза)
и становятся активными только после
поступления в просвет кишки. Трипсиноген
активирует энтерокиназа из клеток
слизистой оболочки кишечника, а также
трипсин. Химотрипсиноген активируется
трипсином, а прокарбоксипептидаза —
карбоксипептидазой.
Липазы. Жиры расщепляет
панкреатическая липаза (гидролизует
триглицериды, ингибитор липазы —
соли жёлчных кислот), холестерол-эстераза
(гидролизует эфиры холестерола) и
фосфолипаза (отщепляет жирные кислоты
от фосфолипидов).
–Амилаза(панкреатическая)
расщепляет крахмал, гликоген и большинство
углеводов до ди- и моносахаридов.
Бикарбонатныеионысекретируют эпителиальные клетки мелких
и средних протоков. Механизм секреции
HCO3–рассмотрен на рис.
22–7,I.
Фазысекрецииподжелудочной
железы такие же, как и желудочной
секреции — мозговая (20% всей секреции),
желудочная (5–10%) и кишечная (75%).
Регуляциясекреции. Секрецию
сока поджелудочной железы стимулируютацетилхолини парасимпатическая стимуляция,холецистокинин,секретин(особенно при очень кислом химусе) ипрогестерон. Действие стимуляторов
секреции обладает эффектом мультипликации,
то есть эффект одновременного действия
всех стимулов намного больше, чем сумма
эффектов каждого стимула по отдельности.
Таблица
22–3.
Нейрогуморальная
регуляция
секреции
холецистокинина
Прогестерон | + |
Блуждающий нерв | + |
Чревные нервы | – |
Эстрогены окситоцинблуждающий нерв | + |
«+» — усиление секреции, «–» —подавление
секреции
Секреция жёлчи
Одной из многообразных функций печени
является желчеобразующая (от 600 до 1000
мл в день). Жёлчь — сложный водный
раствор, состоящий из органических
соединений и неорганических веществ.
Основные компоненты жёлчи — холестерин,
фосфолипиды (главным образом лецитин),
соли жёлчных кислот (холаты), жёлчные
пигменты (билирубин), неорганические
ионы и вода. Жёлчь (первая порция жёлчи)
постоянно секретируется гепатоцитами
и по системе протоков (здесь к жёлчи
добавляется стимулируемая секретином
вторая порция, содержащая много ионов
бикарбоната и натрия) поступает в общий
печёночный и далее в общий жёлчный
проток. Отсюда
печёночная жёлчь опорожняется
непосредственно в двенадцатиперстную
кишку или же поступает в пузырный проток,
ведущий в жёлчный пузырь. Жёлчный пузырь
накапливает и концентрирует жёлчь. Из
жёлчного пузыря концентрированная
жёлчь (пузырная жёлчь) по пузырному и
далее по общему жёлчному протоку порциями
выбрасывается в просвет двенадцатиперстной
кишки. В тонком кишечнике жёлчь участвует
в гидролизе и всасывании жиров.
Концентрированиежёлчи.
Объём жёлчного пузыря — от 30 до 60 мл,
но за 12 часов в жёлчном пузыре может
депонироваться до 450 мл печёночной
жёлчи, так как вода, натрий, хлориды и
другие электролиты постоянно всасываются
через слизистую оболочку пузыря. Основный
механизм абсорбции — активный
транспорт натрия с последующим вторичным
транспортом ионов хлора, воды и других
компонентов. Жёлчь концентрируется в
5 раз, максимально — в 20 раз.
Опорожнениежёлчногопузыряза счёт ритмических сокращений его
стенки происходит тогда, когда пища
(особенно жирная) поступает в
двенадцатиперстную кишку. Эффективное
опорожнение жёлчного пузыря наступает
при одновременном расслаблении сфинктера
Одди. Прием значительного количества
жирной пищи стимулирует полное опорожнение
жёлчного пузыря в течение 1 часа.
Стимулятор опорожнения жёлчного пузыря —
холецистокинин,
дополнительные стимулы поступают
от холинергических волокон блуждающего
нерва.
Функциижёлчныхкислот.
Ежедневно гепатоциты синтезируют около
0,6 г гликохолевых и таурохолевых жёлчных
кислот. Жёлчные кислоты —детергенты,
они уменьшают поверхностное натяжение
жировых частиц, что приводит к
эмульгированию жира. Более того, жёлчные
кислоты способствуют всасыванию жирных
кислот, моноглицеридов, холестерола и
других липидов. Без жёлчных кислот более
40% липидов пищи теряется с калом.
Кишечно–печёночнаяциркуляцияжёлчныхкислот. Жёлчные кислоты
всасываются из тонкой кишки в кровь и
через воротную вену поступают в печень.
Здесь они почти полностью абсорбируются
гепатоцитами и снова секретируются в
жёлчь. Таким способом жёлчные кислоты
циркулируют до 18 раз, прежде чем они
постепенно не будут удалены с калом.
Этот процесс называется кишечно-печёночной
циркуляцией.
Нарушенияобменахолестеринаижёлчныхкислот. Выпадение
холестерина в осадок происходит при
снижении соотношения «холаты/холестерин»
и «лецитин/холестерин», поэтому
образованию камней способствуют
повышение концентрации в жёлчи холестерина
и снижение содержания лецитина и холатов.
Факторы риска: ожирение (повышена
экскреция холестерина с жёлчью),
выраженная гиперлипидемия, нарушение
рационального питания (жирное мясо,
яйца, масло и другие продукты, содержащие
много холестерина), наследственная
предрасположенность, сахарный диабет,
приём ЛС (эстрогены; клофибрат и
никотиновая кислота, используемые для
лечения гиперлипидемии, ингибируют
синтез холестерина и повышают его
экскрецию в жёлчь за счёт мобилизации
из тканей), беременность, резекция тонкой
кишки (нарушается всасывание жёлчных
кислот из-за уменьшения функциональной
поверхности кишечника).
Нарушенияобменабилирубина.
Выделяющийся с жёлчью конъюгированный
билирубин хорошо растворим в воде, но
в ряде случаев, он способен выпадать в
осадок и формировать пигментные камни.
Факторы риска: цирроз печени, хроническая
гемолитическая анемия (перенасыщение
жёлчи билирубином), бактериальная
инвазия жёлчных путей.
Источник
Функции
поджелудочной железы.
I.
Экзокринная. Она заключается
в секреции панкреатического
сока
– смеси
пищеварительных ферментов, поступающих
в двенадцатиперстную кишку и расщепляющих
все компоненты химуса;
II.
Эндокринная.
Она заключается в продукции гормонов.
1 7 2 3 4 6 5 Поджелудочная |
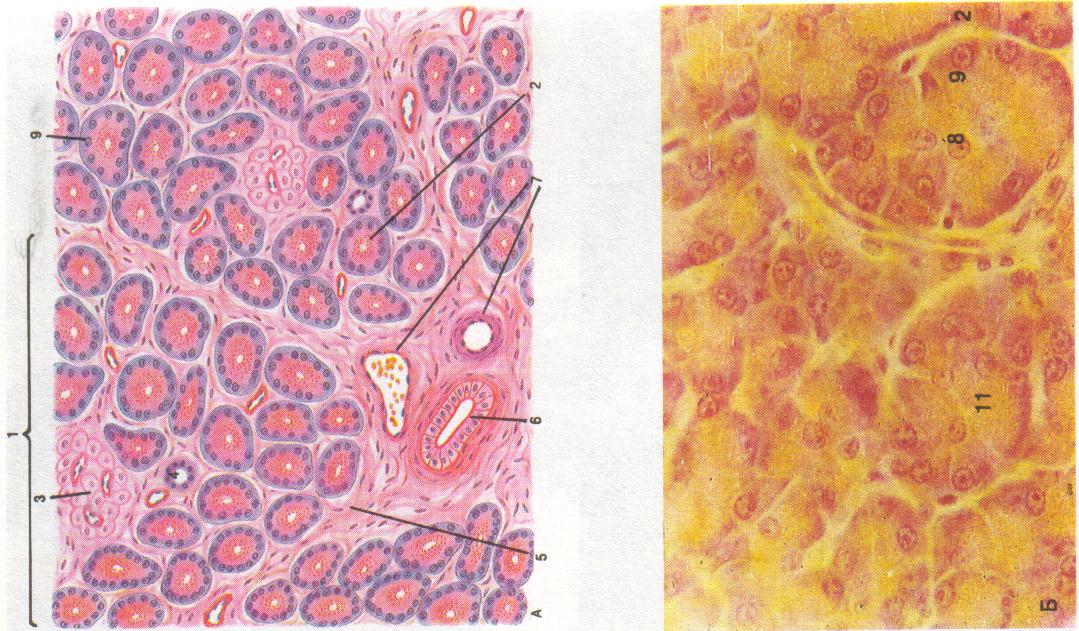
Поджелудочная
железа – паренхиматозный
дольчатый орган.
Строма
железы представлена:
капсулой,
которая сливается с висцеральной
брюшиной и отходя-щими
от неё трабекулами. Строма тонкая,
образована
рыхлой
волокнистой —
тканью. Трабекулы делят железу на дольки.
В прослойках
рыхлой волокнистой ткани находятся
выводные протоки
экзокринной части железы, сосуды, нервы,
интраму-ральные ганглии,
пластинчатые тельца
Фатер-Пачини.
Паренхима
образо-вана совокупностью
секреторных отделов (ацинусов),
выводных протоков и остров-ков
Лангерганса. Каждая
долька состоит из экзокринной и
эндокринной частей. Их соотношение
≈ 97 : 3.
Экзокринная
часть поджелудочной железы представляет
собой сложную
алъвеолярно-трубчатую белковую железу.
Структурно-функциональной
единицей экзокринной части является
панкреатический
ацинус.
Он
образован 8
– 14 ацинозными клетками (ациноцитами)
и центроацинозными клетками
(центроациноцитами).
Ацинозные клетки лежат на базальной
мембране, имеют коническую форму и
выраженную полярность: различающиеся
по строению базальный и апикальный
полюсы. Расширенный базальный полюс
равномерно окрашивается основными
красителями и называется гомогенным.
Суженный апикальный полюс окрашивается
кислыми красителями и называется
зимогенным,
потому
что содержит гранулы зимогена –
проферментов. На апикальном полюсе
ациноцитов имеются микроворсинки.
Функция ациноцитов – выработка
пищеварительных ферментов. Активация
ферментов, секретируемых
ациноцитами, в норме происходит только
в двенадцатиперстной кишке под влиянием
активаторов. Это обстоятельство, а также
вырабатываемые
клетками эпителия протоков ингибиторы
ферментов и
слизь защищают паренхиму поджелудочной
железы от аутолиза (самопереваривания).
Поджелудочная
железа, долька,
рисунок, большое увеличение:
1 – концевой отдел
(ацинус):
а – апикальная
(оксифильная) часть клетки, содержит
зимоген,
б – базальная
(базофильная) – гомогенная часть клетки;
2 – гемокапилляр;
3 – островок
Лангерганса (инсула).
Эндокринная
часть железы. Структурно-функциональной
единицей эндокринной
части поджелудочной железы является
островок
Лангерганса
(инсула). Он
отделён от ацинусов рыхлой волокнистой
неоформленной
тканью. Островок состоит из клеток
инсулоцитов,
между которыми лежит рыхлая волокнистая
соединительная
ткань с гемокапиллярами фенестрированного
типа. Инсулоциты
различаются по способности окрашиваться
красителями. В соответствии
с этим различают инсулоциты типа А, В,
D,
D1,
PP.
В-клетки
(базофильные инсулоциты) окрашиваются
в синий цвет основными красителями. Их
количество составляет около 75% всех
клеток островка. Они располагаются в
центре инсулы. Клетки имеют развитый
белоксинтезирующий аппарат
и секреторные гранулы с широким светлым
ободком. Секреторные
гранулы содержат гормон инсулин
в комплексе с цинком. Функцией
В-инсулоцитов является выработка
инсулина, снижающего в крови
уровень глюкозы и стимулирующего ее
поглощение клетками организма.
В печени инсулин стимулирует образование
из глюкозы гликогена.
[При недостатке выработки инсулина
формируется сахарный
диабет].
А-клетки
(ацидофильные) – составляют 20-25% всех
клеток островка. Они располагаются по
периферии инсулы. Они содержат
гранулы, окрашивающиеся кислыми
красителями. В электронном микроскопе
гранулы имеют узкий ободок. Клетки также
содержат развитый
белоксинтезирующий аппарат и секретируют
гормон глюкагон.
Этот гормон является антагонистом
инсулина (контринсулярный гормон),
поскольку стимулирует распад гликогена
в печени и способствует
повышению содержания глюкозы в крови.
D-клетки
составляют около 5% эндокринных клеток
островка. Они
располагаются по периферии инсулы.
Содержат
умеренно плотные гранулы без светлого
ободка. В гранулах содержится
гормон соматостатин,
угнетающий функцию А, В-клеток островков
и ациноцитов. Он же обладает митозингибирующим
действием
на различные клетки.
D1-клетки
содержат гранулы с узким ободком.
Вырабатывают вазоинтестинальный
полипептид,
понижающий артериальное давление и
стимулирующий выработку панкреатического
сока. Количество этих клеток
невелико.
 РР-клетки
РР-клетки
(2—5%) располагаются по периферии
островков, иногда могут
встречаться и в составе экзокринной
части железы. Содержат гранулы различной
формы, плотности и величины. Клетки
вырабатывают панкреатический
полипептид,
угнетающий внешнесекреторную активность
поджелудочной железы.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Включения — это непостоянные (необязательные) структурные элементы цитоплазмы.
Они заметны при световой микроскопии при общих методах окрашивания, иногда при малом и среднем увеличении, а часть из них можно выявить лишь специальными (гистохимическими, иммунологическими) методами или при электронной микроскопии. В зависимости от активности клетки, гормональных и метаболических влияний, особенностей дифференцировки, возраста, действия разнообразных факторов внешней среды в клетках можно обнаружить большое разнообразие включений по составу и количеству.
Включения указывают на особенности метаболизма, дифференцировки, функциональной активности клеток. Много включений появляется при дистрофических нарушениях в клетке, что сопровождается изменениями в ее жизнедеятельности вплоть до гибели. Иногда содержимое включений не только показатель функции, но основание для названия клетки: пигментные клетки — меланоциты; эозинофильные, базофильные и нейтрофильные гранулоциты крови и др.
При всем многообразии включений их можно объединить по их функциональному назначению.
Секреторные включения. Представляют собой секреторные гранулы, которые выделяются из клетки путем экзоцитоза. По химическому составу их подразделяют на белковые (серозные), жировые (липидные, или липосомы), слизистые (содержат мукополисахариды) и др. Количество включений зависит от функциональной активности клетки, стадии секреторного цикла, степени зрелости клетки. Особенно много гранул в дифференцированных, функционально активных клетках в фазу накопления секреторного цикла.
Секреторные включения образуются в комплексе Гольджи. Перед этим они проходят стадию синтеза в гр. или глад. ЭПС, реже это происходит в других структурах.
Секреторные белковые включения разнообразны по своим размерам, распределению в цитоплазме, электронной плотности. Они окружены клеточной мембраной. Полипептидные цепочки содержимого секреторных включений синтезируются в гр. ЭПС, а созревают в комплексе Гольджи. В связи с этим у клеток, синтезирующих секреторные белки, хорошо развиты данные органеллы, крупное ядро и ядрышки. Однако если клетка прекращает синтез включений, их накопление сопровождается инволюцией гр. ЭПС и комплекса Гольджи.
В экзокринных железах секреторные включения преобладают в апикальной части клетки, предполагая выведение секрета во внешнюю среду. Секреторные включения эндокринных желез концентрируются вблизи кровеносных сосудов или равномерно распределены в цитоплазме.
Слизистые секреторные включения находятся в основном в клетках слизистых секреторных желез. Примером одноклеточных секреторных желез служат бокаловидные клетки тонкой кишки. При световой микроскопии с помощью ШИК-реакции слизь хорошо видна в крупных вакуолях.
Секреторные включения, содержащие жиры (липосомы), имеются в цитоплазме сальных желез и эндокринных клеток, синтезирующих стероидные гормоны (производные холестерина). Стероидные гормоны — это мужские и женские половые гормоны, гормоны стресса (глюкокортикоиды) и гормон, контролирующий содержание ионов натрия в организме (альдостерон). В этих клетках хорошо развита глад, и гр. ЭПС, комплекс Гольджи, много митохондрий. Митохондрии эндокриноцитов участвуют в синтезе стероидных гормонов и имеют специфические особенности строения. Это крупные митохондрии с мультивезикулярными (трубчатыми) кристами.
Также выделяют секреторные включения, содержащие производные аминокислот и других аминов: норадреналин и адреналин, серотонин (мелатонин) и др.
Разнообразен состав секреторных включений в тучной клетке (лаброците) и базофильном гранулоците (базофиле). Эти клетки содержат многочисленные крупные секреторные включения, окрашивающиеся основными красителями и нередко изменяющие их оттенок. Такая способность изменять цвет красителя называется метахромазией. При электронной микроскопии видно, что в лаброцитах и гранулоцитах много крупных гранул округлой формы, различной электронной плотности.
Количество включений зависит от стадии секреторного цикла. Максимально их количество на стадии накопления секрета, а на других стадиях они могут отсутствовать или их концентрация в клетке минимальна.
Трофические включения. Это структуры, в которых клетки и организм в целом запасают питательные вещества, необходимые в условиях энергетического дефицита, недостатка структурных молекул (при голодании). Примером трофических включений служат гранулы с гликогеном (печеночные клетки, мышечные клетки и симпласты), липидные включения в жировых и других клетках.
Трофические включения гликогена представляют собой мелкие, неправильной формы гранулы, которые можно обнаружить при электронной микроскопии, а также при световой микроскопии, используя специальные методы окрашивания. Гликоген при расщеплении превращается в глюкозу, которую используют клетка и организм в целом в условиях ее дефицита.
Липидные включения в норме накапливаются в жировой ткани (белом или буром жире). В липоците белого жира включения сливаются в гигантскую каплю, которая занимает всю центральную часть клетки. Такие клетки приобретают округлую форму, большие размеры. Ядра уплощены и смещены на периферию, органелл немного. В липоцитах бурого жира включения не сливаются в одну каплю, ядра лежат центрально, много митохондрий, развиты комплекс Гольджи и глад. ЭПС.
При переходе на жировой обмен разрушение липидов в жировых тканях поддерживает энергетические потребности организма. Липидные включения легче разрушаются в буром жире, чем в белом. Избыточное накопление липидов в жировой ткани называют ожирением.
Трофические липидные капли могут накапливаться вне жировых клеток: в гепатоцитах, скелетных и сердечных миоцитах, канальцевом аппарате почек и др. Большое накопление таких включений, которое носит обратимый характер и не нарушает функцию клетки, называется жировой инфильтрацией. В случае, когда такое накопление ведет к повреждению клетки, это явление называют жировой дистрофией. Жировая дистрофия стенки артерии — атеросклероз.
Пигментные включения. Этот тип включений придает окраску клеткам; обеспечивает защитную функцию, например, гранулы меланина в пигментных клетках кожи предохраняют от солнечных ожогов. Пигментные включения могут состоять из продуктов жизнедеятельности клетки: гранулы с липофусцином в нейронах, гемосидерин в макрофагах.
Пигментные клетки — меланоциты у низкоорганизованных позвоночных встречаются во многих органах, придавая животным разнообразную окраску. Форма клеток также различная, но в основном многоотростчатая.
У млекопитающих и человека меланоциты встречаются в основном в эпителии. В многослойном эпителии они лежат в базальном слое, а их отростки направляются к шиповатому слою. Пигмент включений меланоцитов — меланин является производным аминокислоты тирозина. Меланин накапливается в многочисленных включениях, располагающихся в теле и отростках клетки. Часть включений выделяется и захватывается соседними клетками. Если клетки не способны вырабатывать меланин, то это ведет к альбинизму.
Экскреторные включения. Это включения веществ, захватываемых клеткой из внутренней среды и выводимых из организма: токсические вещества, продукты метаболизма, инородные структуры. Нередко экскреторные включения встречаются в эпителии канальцев почки, в первую очередь в проксимальных. Проксимальные канальцы выводят ненужные организму вещества, которые не могут быть отфильтрованы через клубочковый аппарат.
Случайные включения. Характерны для фагоцитов, захватывающих чужеродные для организма структуры (частички пыли, бактерии и вирусы), плохо перевариваемые и неперевариваемые макромолекулярные органические и неорганические комплексы. Наиболее часто подобные включения обнаруживают в специализированных клетках, осуществляющих фагоцитоз, — нейтрофильных лейкоцитах и макрофагах.
Минеральные включения. Преимущественно это нерастворимые соли кальция (карбонаты, фосфаты). Они образуются при пониженной активности органа, гипотрофии и атрофии. Нередко минеральные включения (соли кальция) обнаруживают в матриксе митохондрий, это связано с высоким содержанием этого иона и изменением метаболизма в органелле.
Включения при патологии, могут накапливаться в избыточном количестве и вести к нарушению структуры и функции клетки (дистрофии). Дистрофия обусловлена болезнями накопления, связанными с недостаточной активностью лизосом и/или избыточным синтезом каких-либо веществ (жировая дистрофия печени, дистрофия нейронов, при накоплении большого количества гранул с липофусцином, гликогеноз печени и мышц и др.). Накапливаться могут как обычные для клетки вещества (гликоген в гепатоцитах), так и вещества, в норме в клетке не встречающиеся (амилоид).
Большинство включений отделено от матрикса цитоплазмы мембраной (секреторные включения, жировые трофические включения и др.). Однако есть и включения, которые соприкасаются с содержимым гиалоплазмы (гликоген, некоторые минеральные включения).
Происхождение включений разнообразно и зависит от их содержимого. Например, основная масса секреторных и трофических включений формируется в комплексе Гольджи или в ЭПС, а случайные включения, гранулы гемосидерина — продукты неполного переваривания и фагоцитоза.
Утилизация и удаление включений из клетки зависят от природы самого включения. Секреторные включения выводятся из клетки путем экзоцитоза; гликоген и липиды расщепляются ферментами клетки и во внеклеточную среду выводятся в виде продуктов метаболизма (глюкозы, глицерина, жирных кислот); меланин выделяет пигментная клетка, затем его захватывает и разрушает клетка Лангерганса.
Таким образом, включения представляют собой разные по происхождению, функциональному назначению и морфологии структуры. Их число, вид могут быть показателями особенностей дифференцировки и функционального состояния клеток.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Источник

