Внутренняя секреция поджелудочной железы животных
Поджелудочная железа (Pancreas) — железа двойной функции: внешнесекреторной и внутрисекреторной. Внешнесекреторная функция заключается в синтезе и выделении в двенадцатиперстную кишку сока, содержащего пищеварительные ферменты и электролиты, внутрисекреторная — в синтезе и выделении в кровь гормонов.
Внешнесекреторная часть железы сильно развита и составляет более 95 % ее массы. Она имеет дольчатое строение и состоит из альвеол (ацинусов) и выводных протоков. Основная масса ацинусов (железисто-пузырьковидные концевые отделы) представлена панкреатическими клетками — панкреацитами — секретируемыми клетками.
Внутрисекреторная часть железы представлена островками Лангерганса, которые составляют около 30 % массы железы. Различают несколько видов островков Лангерганса по способности секретировать полипептидные гормоны: А-клетки продуцируют глюкогон, В-клетки — инсулин, D-клетки — самостатин. Основную массу островков Лангерганса (около 60 %) составляют В-клетки.
Поджелудочная железа лежит в брыжейке двенадцатиперстной кишки, на печени, разделяясь на правую, левую и среднюю доли. Проток поджелудочной железы открывается в двенадцатиперстную кишку самостоятельно или вместе с желчным протоком. Иногда встречается добавочный проток, который впадает в двенадцатиперстную кишку самостоятельно. Иннервируется поджелудочная железа симпатическими и парасимпатическими нервами (n. vagus).
У собак железа длинная, узкая, красноватого цвета, образует более объемистую левую ветвь и более длинную правую ветвь, достигающую почек. Поджелудочный проток открывается в двенадцатиперстную кишку вместе с желчным протоком. Иногда встречается добавочный проток. Абсолютная масса железы 13-18 г.
У крупного рогатого скота поджелудочная железа располагается вдоль двенадцатиперстной кишки от 12-го грудного до 2-4-го поясничного позвонка, под правой ножкой диафрагмы, частично на лабиринте ободочной кишки. Состоит из поперечной и правой продольной ветвей, соединяющихся под углом в правой стороне. Выводной проток открывается обособленно от желчного протока на расстоянии 30-40 см от него (у овец вместе с желчным протоком). Абсолютная масса железы у крупного рогатого скота 350-500 г, у овец 50-70 г.
У лошадей на поджелудочной железе различают среднюю часть — тело, прилежащее к воротному изгибу двенадцатиперстной кишки. Левый конец железы, или хвост, длинный и узкий, достигает слева слепого мешка желудка, соединяясь с ним, селезенкой и левой почкой. Правый конец железы, или головка, доходит до правой почки, слепой и ободочной кишки. Поджелудочный проток открывается вместе с печеночным. Иногда встречается дополнительный проток. Цвет железы желтоватый, абсолютная масса до 250-350 г.
У свиней на железе различают среднюю, правую и левую доли. Через среднюю долю проходит воротная вена печени. Железа лежит под двумя последними грудными и двумя первыми поясничными позвонками. Проток один, открывается на 13-20 см дистальнее устья желчного протока. Абсолютная масса железы 150 г.
Внешнесекреторная (экзогенная) функция поджелудочной железы. Основной продукт внешнесекреторной функции поджелудочной железы — пищеварительный сок, который содержит 90 % воды и 10% плотного осадка. Плотность сока 1,008-1,010; рН 7,2-8,0 (у лошадей 7,30-7,58; у крупного рогатого скота 8). В состав плотного осадка входят белковые вещества и минеральные соединения: бикарбонат натрия, хлорид натрия, хлорид кальция, фосфорнокислый натрий и др.
Сок поджелудочной железы содержит протеолитические и нуклеолитические ферменты (трипсин, хемотрипсин, карбоксипептидазы, эластазу, нуклеазы, аминопептидазу, коллагеназу, дипептидазу), амилолитические ферменты (а-амилазу, мальтазу, лактазу, инвертазу) и липолитические ферменты (липазу, фосфолипазу, холинэстеразу, карбоксиэстеразу, моноглицеридлипазу, щелочную фосфатазу). Трипсин расщепляет белки до аминокислот и выделяется в виде неактивного трипсиногена, который активируется ферментом кишечного сока энтерокиназой. Химотрипсин расщепляет белки и полипептиды до аминокислот и выделяется в форме неактивного химотрипсиногена; активируется трипсином. Карбоксиполипептидазы действуют на полипептиды, отщепляя от них аминокислоты. Дипептидазы расщепляют дипептиды на свободные аминокислоты. Эластаза действует на белки соединительной ткани — эластин, коллаген. Протаминаза расщепляет протамины, нуклеазы — нуклеиновые кислоты на мононуклеотиды и фосфорную кислоту.
При воспалении поджелудочной железы, аутоиммунных процессах протеолитические ферменты становятся активными уже в самой железе, вызывая ее разрушение. а-Амилаза расщепляет крахмал и гликоген до мальтозы; мальтаза — мальтозу до глюкозы; лактаза расщепляет молочный сахар на глюкозу и галактозу (она имеет существенное значение в пищеварении молодняка), инвертаза — сахарозу на глюкозу и фруктозу; липаза и другие липолитические ферменты расщепляют жиры на глицерин и жирные кислоты. Липолитические ферменты, в частности липаза, секретируются в активном состоянии, но расщепляют только жир, эмульгированный желчными кислотами. Амилазы, также как и липазы, в соке поджелудочной железы находятся в активном состоянии.
Из электролитов в соке поджелудочной железы содержатся натрий, калий, хлор, кальций, магний, цинк, медь и значительное количество бикарбонатов, обеспечивающих нейтрализацию кислого содержимого двенадцатиперстной кишки. Тем самым создается оптимальная среда для активных ферментов.
Доказано, что помимо перечисленного выше действия сок поджелудочной железы обладает свойством регуляции микробной ассоциации в двенадцатиперстной кишке, оказывая определенное бактерицидное действие. Прекращение поступления в кишечник панкреатического сока ведет к усиленному бактериальному росту в проксимальном отделе тонкого кишечника у собак.
Эндокринная (гормональная) функция поджелудочной железы. Важнейшими гормонами поджелудочной железы являются инсулин, глюкогон и соматостатин.
Инсулин образуется в В-клетках из предшественника — проинсулина. Синтезируемый проинсулин поступает в аппарат Гольджи, где расщепляется на молекулу С-пептида и молекулу инсулина. Из аппарата Гольджи (пластинчатый комплекс) инсулин, С-пептид и частично проинсулин поступают в везикулы, где инсулин связывается с цинком и депонируется в таком состоянии. Под влиянием различных стимулов инсулин освобождается от цинка и поступает в прекапиллярное пространство. Основным стимулятором секреции инсулина служит глюкоза: при ее повышении в крови усиливается синтез инсулина. В определенной степени этим свойством обладают аминокислоты аргинин и лейцин, а также глюкогон, глетрин, секретин, глюкокортикоиды, соматостатин, никотиновая кислота. Инсулин в крови находится в свободном и связанном с белками плазмы состоянии. Распад инсулина происходит в печени под влиянием глютатионтрансферазы и глютатионредуктазы, в почках под влиянием инсулиназы, в жировой ткани под влиянием протеолитических ферментов. Проинсулин и С-пептид тоже подвергаются дегидратации в печени. Его биологическое действие обусловлено способностью связываться со специфическими рецепторами клеточной цитоплазматической мембраны.
Инсулин усиливает синтез углеводов, белков, нуклеиновых кислот и жира. Он ускоряет транспорт глюкозы в клетки инсулинозависимых тканей (печень, мышцы, жировая ткань), стимулирует синтез гликогена в печени и подавляет глюконеогенез (образование глюкозы из неуглеводных компонентов), гликогенолиз (распад гликогена), что в конечном итоге ведет к снижению уровня сахара в крови. Этот гормон ускоряет транспорт аминокислот через цитоплазматическую мембрану клеток, стимулирует синтез белка. Инсулин участвует в процессе включения жирных кислот в триглицериды жировой ткани, стимулирует синтез липидов и подавляет липолиз (распад жира).
В регуляции синтеза белка и утилизации углеводов вместе с инсулином участвуют кальций и магний. Концентрация инсулина в крови человека 15-20 мкЕД/мл.
Глюкогон — полипептид, секреция которого регулируется глюкозой, аминокислотами, гастроинтестинальными гормонами (панклеозимин) и симпатической нервной системой. Секреция глюкогона усиливается при снижении в крови сахара, СЖК, раздражении симпатической нервной системы, а угнетается при гипергликемии, повышении уровня СЖК, соматостатина. Под влиянием глюкогона стимулируется глюконеогенез, ускоряется распад гликогена, т. е. увеличивается продукция глюкозы. Под действием глюкогона ускоряется синтез активной формы фосфорилазы, участвующей в образовании глюкозы из неуглеводных компонентов (глюконеогенез). Глюкогон способен связываться с рецепторами адипацитов (клеток жировой ткани), способствуя распаду триглицеридов с образованием глицерина и СЖК. Глюконеогенез сопровождается не только образованием глюкозы, но и промежуточных продуктов обмена веществ — кетоновых тел, развитием кетоацидоза. Содержание в плазме крови глюкогона у человека составляет 50-70пг/мл. Концентрация этого гормона в крови увеличивается при голодании (голодный кетоз у овец), хронических заболеваниях печени.
Соматостатин — гормон, основной синтез которого осуществляется в гипоталамусе, а также в D-клетках поджелудочной железы. Соматостатин подавляет секрецию СТГ, АКТГ, ТТГ, гастрина, глюкогона, инсулина, ренина, секретина, вазоактивного желудочного пептида, желудочного сока, панкреатических ферментов и электролитов. Содержание соматостатина в крови повышается при сахарном диабете I типа, D-клеточной опухоли поджелудочной железы (соматостатиноме). Говоря о гормонах поджелудочной железы, следует отметить, что энергетический баланс в организме поддерживается сплошными биохимическими процессами, в которых непосредственное участие принимают инсулин, глюкогон и частично соматостатин. Так, во время голодания уровень в крови инсулина снижается, а глюкогона повышается, усиливается глюконеогенез. Благодаря этому поддерживается минимальный уровень глюкозы в крови. Усиление липолиза сопровождается повышением в крови СЖК, которые используются сердечной и другими мышцами, печенью, почками в качестве энергетического материала. В условиях гипогликемии источником энергии становятся и кетокислоты.
Нейроэндокринная регуляция функции поджелудочной железы. Деятельность поджелудочной железы находится под влиянием парасимпатической (n. vagus) и симпатической (чревные нервы) нервной системы, гипоталамогипофизарной системы и других желез внутренней секреции. В частности, блуждающий нерв играет определенную роль в регуляции ферментообразования. Секреторные волокна входят также в состав симпатических нервов, иннервирующих поджелудочную железу. При стимуляции отдельных волокон блуждающего нерва с усилением сокоотделения происходит и его торможение. Основоположником отечественной физиологии И. П. Павловым доказано, что отделение поджелудочного сока начинается при виде корма или раздражении рецепторов полости рта и глотки. Этот феномен необходимо учитывать в случаях назначения голодной диеты при остром панкреатите у собак, кошек и других животных, не допуская их зрительного и обонятельного контакта с кормом.
Наряду с нервной происходит и гуморальная регуляция функции поджелудочной железы. Поступление соляной кислоты в двенадцатиперстную кишку вызывает секрецию поджелудочного сока даже после перерезки блуждающих и чревных (симпатических) нервов и разрушения продолговатого мозга. Это положение лежит в основе назначения медикаментов, снижающих секрецию поджелудочного сока при остром панкреатите. Под влиянием соляной кислоты желудочного сока, поступающего в кишечник, из клеток слизистой оболочки тонкой кишки выделяется просекретин. Соляная кислота активирует просекретин, превращая его в секретин. Всасываясь в кровь, секретин действует на поджелудочную железу, усиливая выделение ею сока: одновременно он тормозит функцию обкладочных желез, чем препятствует чрезмерно интенсивной секреции соляной кислоты железами желудка. Секретин в физиологическом отношении является гормоном. Под влиянием секретина образуется большое количество поджелудочного сока, бедного ферментами и богатого щелочами. Учитывая эту физиологическую особенность, лечение острого панкреатита направлено на снижение секреции соляной кислоты в желудке, подавление активности секретина.
В слизистой оболочке двенадцатиперстной кишки образуется также гормон панкреозимин, который усиливает образование ферментов в поджелудочном соке. Подобное действие оказывают гастрин (образуется в желудке), инсулин, соли желчных кислот.
Тормозящее влияние на секрецию панкреатического сока оказывают нейропептиды — гастроингибирующий полипептид (ГИП), панкреатический полипептид (ПП), вазоактивный интерстинальный полипептид (ВИП), а также гормон соматостатин.
При лечении плотоядных животных с нарушением внешнесекреторной функции поджелудочной железы необходимо иметь в виду, что на молоко выделяется мало сока, на мясо, черный хлеб — много. При кормлении мясом выделяется много трипсина, при кормлении молоком — много липазы и трипсина.
Источник
желез
обусловлены нарушением состояния ЦНС.
После перерезки двигательных нервов
судороги денервированных мышц не
возникают.
Паратиреопривная
тетания развивается вследствие
понижения уровня кальция в крови и
спинномозговой жидкости. Введение
солей кальция таким животным предупреждает
развитие тетании. При тетании нарушаются
также функции печени; в крови обнаруживается
токсичный карбаминово-кислый аммоний.
При
недостаточности внутрисекреторной
функции околощитовидных желез у человека
(гипопаратиреоз) вследствие падения
уровня кальция в крови резко повышается
возбудимость ЦНС и возникают приступы
судорог. При скрытой тетании, возникающей
при легкой недостаточности околощитовидных
желез, судороги мышц лица и рук появляются
только при надавливании на нерв,
иннервирующий эти мышцы.
У
детей с врожденной недостаточностью
паращитовидных желез содержание кальция
в крови снижено, нарушен рост костей,
зубов и волос, наблюдаются длительные
сокращения мышечных групп (предплечья,
грудной клетки, глотки и др.).
Избыточная
функция (гиперпарати-реоз) околощитовидных
желез наблюдается довольно редко,
например при опухоли околощитовидной
железы. При этом содержание кальция
в крови увеличено, а количество
неорганического фосфата уменьшено.
Развивается остеопороз, т. е. разрушение
костной ткани, мышечная слабость
(вынуждающая больного постоянно лежать),
боли в спине, ногах и руках. Своевременное
удаление опухоли восстанавливает
нормальное состояние.
Околощитовидные
железы продуцируют паратгормон.
При недостатке паратгормона понижается,
а при избытке повышается содержание
кальция в крови. Одновременно в первом
случае увеличивается содержание в крови
фосфатов и уменьшается их выделение
с мочой, а во втором случае
—
понижается количество фосфатов в крови
и повышается их выделение с мочой.
Паратгормон активирует функцию
остеокластов, разрушающих костную
ткань.
В
организме паратгормон вызывает разрушение
костной ткани с выходом из нее ионов
кальция (вследствие чего и повышается
их концентрация в крови). Паратгормон
усиливает всасывание кальция в кишечнике
и процессы его реабсорбции в канальцах
почди. Все это ведет к значительному
нарастанию уровня кальция в крови
(вместо нормальных
9—11
мг% до
18
мг% и выше). Одновременно снижается
концентрация неорганических фосфатов
в крови и увеличивается их выделение с
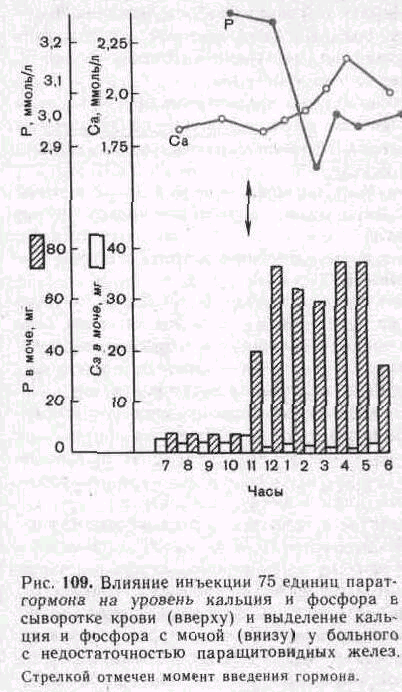
мочой (рис.
109).
В
норме концентрация ионов
Ca2’1′
в плазме крови поддерживается на
постоянном уровне, являясь одним из
наиболее точно регулируемых параметров
внутренней среды. Падение уровня кальция
в крови, омывающей железу, приводит к
усилению секреции паратгормона и,
следовательно, к увеличению поступления
кальция в кровь из его костных депо.
Наоборот, повышение содержания этого
электролита в крови, омывающей
паращитовидные железы, непосредственно
угнетает выделение паратгормона (и
усиливает образование тирокальцитонина),
в результате чего количество кальция
в крови снижается. Таким образом,
между содержанием кальция в крови и
внутренней секрецией околощитовидных
желез (и парафолликулярных клеток
щитовидной железы) имеется непосредственная
двусторонняя связь: смещение концентрации
кальция в омывающей их крови вызывает
изменения секреции тирокальцитонина
и паратгормона, а последние регулируют
содержание кальция в крови.
Указанные
реакции железы на изменение содержания
Са24″
в крови не опосредованы какими-либо
нервными или гуморальными механизмами.
Они являются прямыми и возникают не
только в целом организме, но и при
перфузии изолированной железы кровью,
содержащей .большее или меньшее по
сравнению с нормой количество кальция.
Гистологическими
исследованиями поджелудочной железы
установлено, что в ней наряду с секреторным
эпителием, выделяющим пищеварительные
ферменты, существуют особые группы
клеток
—
белые отростчатые эпидермоциты (островки
Лангерганса
— по
имени открывшего их исследователя).
Эти эпидермоциты не имеют выводных
прртоков и выделяют свой секрет
непосредственно в кровь.
Еще
в конце
XIX
в. было установлено, что у собаки через
4—5
ч после удаления поджелудочной железы
начинается выделение сахара с мочой.
Резко повышается содержание глюкозы
в крови. Потеря сахара с мочой приводит
к тому, что животное худеет, пьет
много воды, становится прожорливым.
Все
эти явления оказались аналогичны тем,,
которые наблюдаются у человека при
сахарном диабете. После пересадки
животному поджелудочной железы в
какой-либо другой участок тела,
например под кожу, проявления сахарного
диабета исчезали.
Для
сахарного диабета характерно повышение
содержания глюкозы в крови (гипер-гликемия)
до
10
ммоль/л
(200
мг%) и даже больше, вместо 4,4±1,1 ммоль/л
(100— 120
мг%) в норме. Это связано с тем, что при
диабете поступившая в кровь глюкоза не
полностью утилизируется тканями и не
превращается в гликоген печени.
Повышение
содержания глюкозы в крови, а следовательно,
и в клубочковом фильтрате приводит
к тому, что эпителий почечных канальцев
не реабсорбирует глюкозу полностью,
вследствие чего она выделяется с мочой
(глюкозурия). Возникает потеря сахара
с мочой
—
сахарное мочеизнурение.
Количество
мочи увеличено (полиурия). Причина этого
явления заключается в том, что при
большом содержании глюкозы в моче
почечных канальцев эта нереабсорбиро-ванная
глюкоза, создавая высокое осмотическое
давление мочи, удерживает в ней воду.
Последняя недостаточно всасывается
канальцами, и количество выделяемой
почками мочи оказывается увеличенным.
Обеднение организма водой вызывает у
больных диабетом сильную жажду, что
приводит к обильному приему воды
(полидипсия). В связи с выведением глюкозы
с мочой резко увеличивается расходование
белков и жиров в качестве веществ,
обеспечивающих энергетический обмен
организма. Об усилении процессов
сгорания жиров и белков свидетельствует
снижение дыхательного коэффициента
нередко до
0,7.
В
организме накапливаются продукты
неполного окисления жиров, к числу
которых относятся кетоновые тела:
Р-оксимасляная и ацетоуксусная кислоты.
В
тяжелых случаях интенсивное образование
кислых продуктов расщепления жиров и
дезаминирование аминокислот в печени
вызывают сдвиг активной реакции крови
в кислую сторону
—
ацидоз.
Накопление
кетокислот и ацидоз могут вызывать
тяжелое, угрожающее смертью состояние
—
диабетическую кому, которая протекает
с потерей сознания, нарушением дыхания
и кровообращения.
Описанные
расстройства связаны со снижением
гормональной функции поджелудочной
железы.
Гормоны поджелудочной железы
Белые
отростчатые эпидермоциты (островки
Лангерганса) состоят из клеток трех
типов: а-, |3- и ^-клеток. Среди них больше
всего (З-клеток (у собак около
75 %);
они небольших размеров и имеют
зернистую протоплазму.
Бета-клетки
выделяют инсулин
(от латинского слова
insula—островок).
Альфа-клетки островков вырабатывают
гормон глюкагон.
По
данным некоторых авторов, эпителии
мелких протоков поджелудочной железы
выделяет гормон липокаин. В экстрактах
этой железы найдены еще два гормона
— ваготонин
и центропнеин.
Инсулин.
Попытки извлечь из поджелудочной железы
инсулин долгое время оставались
тщетными, так как этот гормон является
полипептидом и разрушается трипсином,
содержащимся в ткани вырезанной из
организма поджелудочной железы.
В
1902
г. Л. В. Соболев предложил два способа,
позволяющих предотвратить разрушение
инсулина. Один из этих способов
состоит в том, что у животного перевязывают
протоки поджелудочной железы за несколько
дней до ее удаления. Это вызывает
дегенерацию и гибель внешнесекретор-ного
эпителия. Вследствие этого в железе не
содержится более сока, который мог бы
вызвать ферментативное расщепление
инсулина. Второй способ состоит в том,
что инсулин получают из поджелудочной
железы эмбрионов, в которой еще не
образуется пищеварительных ферментов.
В
1922
г. Бантинг и Бест, применив первый из
этих способов, получили активные
препараты инсулина. Инсулин является
лечебным средством при диабете. Он
поддерживает жизнь не менее чем
30
млн. живущих на планете больных
диабетом, причем
30—40%
из них нуждаются в постоянном ежедневном
введении инсулина.
Инсулин
(полипептид) удалось синтезировать
химическим путем. Это был первый белок,
полученный синтетически вне организма.
Инсулин, полученный из поджелудочной
железы разных видов животных, различается
расположением аминокислот в молекуле.
Молекула инсулина не содержит цинка,
однако способна связывать цинк; при
этом эффект действия инсулина удлиняется
и усиливается.
Инсулин
резко повышает проницаемость мембраны
мышечных и жировых клеток для глюкозы.
Вследствие этого скорость перехода
глюкозы внутрь этих клеток увеличивается
примерно в
20
раз по сравнению со скоростью перехода
глюкозы в клетки в среде, не содержащей
инсулина.
Ферментативные
реакции, приводящие к утилизации
глюкозы,—.
фосфорилирование и окисление ее, а также
образование гликогена протекают внутри
клетки. Способствуя транспорту глюкозы
внутрь клетки, инсулин тем самым
обеспечивает ее утилизацию. Вместе с
тем он не оказывает влияния на утилизацию
углеводов бесклеточными гомоге-натами
тканей (гомогенаты получают путем
растирания клеток, при котором разрушаются
клеточные мембраны), так как механизм
влияния инсулина на углеводный обмен
связан именно с действием его на
проницаемость клеточной мембраны.
Увеличение
транспорта глюкозы через мембраны
мышечных волокон при действии инсулина
способствует синтезу гликогена и
накоплению его в мышечных волокнах. В
клетках жировой ткани инсулин стимулирует
образование жира из глюкозы.
Под
влиянием инсулина возрастает проницаемость
клеточной мембраны и для аминокислот,
из которых в клетках синтезируются
белки. Инсулин стимулирует синтез
информационной РНК и этим также
способствует синтезу белков.
Мембраны
клеток печени в отличие от мембраны
клеток жировой ткани и мышечных
волокон свободно проницаемы для глюкозы
и в отсутствие инсулина. Предполагают,
что этот гормон действует непосредственно
на углеводный обмен печеночных клеток,
активируя синтез гликогена.
Возникающий
после введения больших доз инсулина
переход значительного количества
глюкозы из плазмы крови внутрь клеток
скелетной мускулатуры, сердечной мышцы,
гладких мышц, молочной железы и некоторых
других /органов вызывает падение уровня
глюкозы в крови и вследствие этого
недостаточное поступление глюкозы в
клетки
нервной системы (на проницаемость
которых инсулин не действует). Поэтому
головной и спинной мозг начинает
испытывать острый недостаток глюкозы,
которая является основным источником
энергии для нервных клеток. Когда
содержание сахара в крови падает до
2,5
ммоль/л
(45—50
мг%) возникает острое нарушение
деятельности мозга
— гипогликемическая
кома. Появляются периодические приступы
судорог, затем падение мышечного тонуса,
понижение температуры тела, потеря
сознания. Гипогликемическая кома может
возникать даже под влиянием небольшой
дозы инсулина, если он вводится натощак,
когда глюкоза из пищеварительного
тракта в кровь не поступает. Внутривенное
введение раствора глюкозы немедленно
купирует гипогликемическую кому.
Глюкагон.
Второй гормон поджелудочной железы
—
глюкагон
—
выделяется а-клетками белых отростчатых
эпидермоцитов. Глюкагон стимулирует
внутри клетки переход неактивной
фосфорилазы (фермента, принимающего
участие в расщеплении гликогена с
образованием глюкозы) в активную форму
и тем самым усиливает расщепление
гликогена (в печени, но не в мышцах),
повышая уровень сахара в крови.
Одновременно глюкагон стимулирует
синтез гликогена в печени из аминокислот.
Глюкагон тормозит синтез жирных
кислот в печени, но активирует печеночную
липазу, способствуя расщеплению
жиров. Он стимулирует также расщепление
жира в жировой ткани. Глюкагон повышает
сократительную функцию миокарда, не
влияя на его возбудимость.
Источник

