
Железистый эпителий поджелудочной железы
ПЖ состоит преимущественно из экзокринной ткани. Основной элемент экзокринной части ПЖ — ацинусы: они вместе с разветвлённой сетью протоков составляют 75—90% массы железы. Аиинусы представляют собой субъединицы долек ПЖ и состоят из пирамидальных клеток, обращенных апикальной частью к секреторному канальцу (см. рис. 1-8).

Рис. 1-8. Нормальная гистология поджелудочной железы, микрофотографии: а — малое увеличение; показано соотношение между экзокринной паренхимой поджелудочной железы, протоковой системой и островками Лангерганса (окраска гематоксилином и эозином, увеличение ×75); б — при большем увеличении показаны основные структурные элементы поджелудочной железы, хорошо видны островки и ацинусы (окраска гематоксилином и эозином, увеличение ×100); в — большое увеличение; обнаруживаются заметные различия между экзо- и эндокриноцитами
Секреторные канальцы ацинусов, сливаясь между собой, образуют внутридольковые протоки.
Экзокринная ткань ПЖ состоит из трёх типов клеток:
• ацинарных, продуцирующих гликолитические, липолитические и протеолитические ферменты (в неактивной форме: в виде проферментов или зимогенов) и составляющих до 80% клеточного состава ПЖ;
• центроацинарно-дуктулярных, секретирующих содержащую бикарбонаты жидкость;
• муцинсекретирующих протоковых.
Ацинарные клетки — основной структурный компонент ацинусов и ПЖ в целом. Ациноцит имеет форму усечённого конуса, широкое основание клетки называют базальным отделом, а противоположную, суженную и обращенную в проток часть клетки — апикальной частью. Апикальная часть клетки имеет множество микроворсинок. Ациноииты синтезируют и выделяют в полость ацинуса белковый секрет, 98% которого составляют ферменты.
От полости ацинуса — центроацинарного протока, образованного апикальными поверхностями ашшоцитов, — начинается вставочный отдел панкреатического протока, стенка которого образована мелкими центроацинарными клетками (плоский эпителий). За вставочным отделом протоков лежат межацинарный и внутридольковый протоки, отводящие секрет от первичной дольют железы. За ними следуют остальные порядки протоковой системы: междольковые, междолевые и главные выводные протоки, все вместе образующие выводную (протоковую) систему ПЖ.
Главные и междольковые протоки выстланы высоким призматическим эпителием, внутридольковые — кубическим. Эпителий представлен протоковыми клетками, на долю которых приходится до 5% клеточной массы ПЖ.
Базолатеральные ацинарные клетки (см. рис. 1-9 a) имеют хорошо развитую грубую эндоплазматическую сеть, в которой происходит синтез панкреатических ферментов. После синтеза зимогены попадают в комплекс Гольджи, где происходит их сортировка с другими клеточными белками, а затем в сосуды накопления. В этих сосудах формируются зимогенные гранулы, перемещающиеся к апикальной части клетки (см. рис. 1-9 б), При стимуляции ациноциты выбрасывают содержимое гранул в просвет ацинуса, и секрет поступает в просвет кишечника через панкреатический проток.

Рис. 1-9. Панкреатические ацинарные клетки, электронные микрофотографии
Эндокринная часть ПЖ состоит из небольших островков, известных как островки Лангерганса (см, рис. 1-10). Они отделены от ацинусов прослойками соединительной ткани, густо васкуляризированы, не имеют выводных протоков и содержат следующие типы клеток:
• а-клетки, секретирующие глюкагон, пептид YY;
• в-клетки, секретирующие инсулин, С-пептид, панкреастатин;
• D-клетки, секретирующие соматостатин;
• РР- (или F-) клетки, секретирующие панкреатический полипептид.

Рис. 1-10. Островок Лангерганса, электронные микрофотографии: а — α-клетки; б — β-клетки; в — D-клетки; увеличение ×240
в-Клетки наиболее многочисленны и расположены в центре островков. Соотношение a-, D- и F-клеток, лежащих по периферии островков, неодинаково в каждом ацинусе. Передняя часть железы содержит больше F-клеток, тогда как в задней части больше в-клеток. Физиологическое значение таких региональных различий до конца не изучено, но наличие клеток разных типов необходимо для паракринной регуляции функции островков Лангерганса с помощью соматостатина. Соматостатин, в свою очередь, регулирует высвобождение других гормонов — инсулина и глюкагона.
Маев И.В., Кучерявый Ю.А.
Опубликовал Константин Моканов
Источник
Железы представляют собой органы, которые синтезируют и выделяют различные вещества, необходимые для жизнедеятельности организма. Большинство желез образовано эпителиальной тканью – железистым эпителием. Исключение составляют мозговой слой надпочечников, задняя доля гипофиза и эпифиз, паренхима которых имеет нейрогенное происхождение.
Железистые клетки – гландулоциты — являются высокоспециализированными клетками, которые специализируются на синтезе, накоплении и выведении секрета. Ядро гландулоцитов, как правило, крупное, имеет одно или несколько ядрышек. Цитоплазма содержит многочисленные органоиды; особенно хорошо развит пластинчатый комплекс. Характер развития органоидов зависит от химической природы синтезируемых веществ. Распределение органоидов в цитоплазме неравномерно, клетки отличаются выраженной полярностью. Процесс синтеза, накопления и выделения секрета гландулоцитом обозначается как секреторный цикл. Он состоит из четырех фаз: поглощения исходных веществ, синтеза компонентов, накопления продукта и его выведения. Фаза поглощения обеспечивается транспортными системами, которые связаны с плазмолеммой базальной части клетки. В большинстве случаев исходные вещества поступают непосредственно из крови, но иногда они могут запасаться в цитоплазме гландулоцита. Фаза синтеза связана с деятельностью шероховатой и гладкой плазматической сети, пластинчатого комплекса и митохондрий. Синтезированный продукт накапливается в пластинчатом комплексе, где происходит его созревание и упаковка в одномембранные пузырьки. Фаза накопления состоит в появлении в цитоплазме гландулоцита секреторных гранул, которые укрупняются, сливаясь между собой. Фаза выведения обычно заключается в экзоцитозе секреторных гранул или вакуолей (экструзии), однако продукты могут выводиться и диффузно.
Наиболее короткой является фаза поглощения, она протекает не более 5 мин. Продолжительность фаз синтеза и накопления составляет около 10-15 мин. Самой продолжительной является фаза выведения, которая занимает 30 мин. Общее время секреторного цикла составляет около одного часа.
Железистый эпителий образует органы двух типов:
· экзокринные железы, которые выделяют секрет на поверхность тела или в просвет внутренних органов;
· эндокринные железы, которые выводят продукт (инкрет, или гормон) в кровь.
Экзокринные железыимеютконцевые (секреторные) отделы и выводные протоки. Концевой отдел (ацинус) экзокринной железы состоит из железистых клеток, которые в один или несколько слоев располагаются на базальной пластинке и продуцируют секрет. Выводной проток также состоит из эпителиальных клеток, растущих на базальной пластинке, но они обычно не секретируют, а обеспечивают связь концевого отдела с покровным эпителием.
В морфологической классификации экзокринных желез используются такие их признаки как форма концевых отделов (трубчатая, альвеолярная, трубчато-альвеолярная), ветвление концевого отдела (разветвленный и неразветвленный), ветвление выводного протока (простая и сложная железы).
Морфологическая классификация экзокринных желез
| Железа | Тип по морфологической классификации |
| Сальная железа | Простая неразветвленная альвеолярная |
| Потовая железа | Простые неразветвленная трубчатые |
| Слюнная железа | Сложная разветвленная альвеолярная |
| Поджелудочная железа | Сложная альвеолярная |
| Молочная железа | Сложная альвеолярно-трубчатая |
Кроме морфологической классификации экзокринные железы подразделяют также на группы в зависимости от количества клеток (одноклеточные, малоклеточные и многоклеточные), расположения относительно эпителиального пласта (экзоэпителиальные и эндоэпителиальные), состава секрета (белковые, слизистые, белково-слизистые, липидные, кислотные). Экзокринные железы классифицируют также по типу секреции:
1. Голокриновый тип, при котором происходит гибель и разрушение гландулоцита (сальная железа).
2. Макроапокриновый тип, при котором наблюдается утрата материала апикального конца клетки (молочная железа).
3. Микроапокриновый тип, который отличается отрывом расширенных вершин микроворсинок (хориоидное сплетение в третьем желудочке мозга).
4. Мерокриновый (эккриновый) тип, при котором не наблюдается явных изменений морфологии гландулоцитов (большинство желез).
В качестве примера приведем краткую морфо-функциональную характеристику сальной, поджелудочной (экзокринная часть) и молочной желез.
Сальная железа находится рядом с волосом. Она состоит из небольшого количества расположенных на базальной пластинке клеток, которые делятся, синтезируют секрет и разрушаются. Ее единственный проток впадает во влагалище волоса. Клеточный детрит содержит жироподобный секрет, необходимый для смазывания роговых чешуй. Тип секреции – голокриновый.
Экзокринная часть поджелудочной железы состоит из многочисленных долек, стенки которых образованы крупными клетками однослойного эпителия, расположенного на базальной пластинке. В базальной части гландулоцита поджелудочной железы находится большинство органоидов, тогда как в апикальной части локализованы гранулы зимогена (комплекса пищеварительных ферментов). Гранулы путем экструзии выходят в просвет дольки и по протокам достигают кишечника. Тип секреции – мерокриновый.
Молочная железа выделяет секрет сложного состава, который содержит много липидов, специфических белков, сахаров и неорганических веществ, особенно кальция. Она состоит из тяжей эпителиальных клеток, располагающихся между прослойками соединительной ткани. Гландулоциты молочной железы крупные, с большим ядром, развитой плазматической сетью и пластинчатым комплексом. Во время лактации от апикальной части гландулоцитов отрываются пузырьки различного размера. Тип секреции – макроапокриновый.
Эндокринные железы в отличие от экзокринных не имеют выводных протоков, представляя собой группы эпителиальных клеток, оплетенных кровеносными капиллярами. Эндокринные железы синтезируют и выделяют в кровь гормоны (инкреты), которые представляют собой биологически активные вещества, регулирующие многие жизненно важные функции организма.
Эндокринные железы имеют различное строение. Они могут быть представлены отдельными клетками (например, энтерохромаффинные клетки кишечного эпителия, которые в совокупности образуют диффузную эндокринную систему) или органами различной величины (гипофиз, надпочечники, щитовидная железа). Особенности гистофизиологии эндокринных желез можно рассмотреть на примере трех органов: передней доли гипофиза, островков поджелудочной железы и щитовидной железы.
Гипофиз – это эндокринная железа шаровидной формы, которая связывает головной мозг и эндокринные железы в единую систему. Он состоит из трех долей: передней, промежуточной и задней. Передняядоля(аденогипофиз) состоит из соединительно-тканной оболочки и тесно прилежащих эпителиальных клеток, которые оплетены кровеносными капиллярами. При окраске гематоксилин-эозином в аденогипофизе выделяют три типа клеток: оксифильные, базофильные и хромофобные. Оксифильные и базофильные клетки синтезируют и накапливают пептидные гормоны, которые концентрируются в секреторных гранулах диаметром 20-100 нм. При этом каждая клетка секретирует только один тип гормона. Хромофобные клетки не содержат гранул с гормонами и находятся вне секреторного цикла.
Аденогипофиз координирует деятельность всех других эндокринных желез. Он секретирует в кровь такие гормоны как соматотропин (СТГ, или гормон роста – усиливает анаболические процессы), аденокортикотропин (АКТГ — усиливает катаболические процессы), тиреотропин (ТТГ – контролирует щитовидную железу), лютеинизирующий и фолликулостимулирующий гормоны (ЛГ и ФСГ – регулируют созревание яйцеклеток и сперматозоидов) и ряд других. Нарушения функций клеток аденогипофиза приводят к тяжелым формам гормональных дисфункций – карликовости, умственной отсталости, бесплодию у мужчин и женщин.
Панкреатические островки (островки Лангерганса-Соболева) представляют собой небольшие группы клеток, расположенные около сосудов между дольками экзокринной части. Клетки панкреатических островков синтезируют и выделяют гормоны, регулирующие уровень сахара в крови. Как и в аденогипофизе, каждая клетка островка секретирует только один вид гормона, который упаковывается в гранулы. В панкреатическом островке различают четыре типа клеток: А, В, С и D. А — клетки синтезируют глюкагон, который способствует образованию глюкозы из гликогена. В — клетки синтезируют инсулин, стимулирующий превращение глюкозы в гликоген. D — клетки выделяют соматостатин, снижающий уровень гормона роста в крови. С — клетки гранул не имеют, они являются предшественниками В-клеток. Для медицины особенно важны генетические дефекты В-клеток панкреатических островков, приводящие к недостатку инсулина и развитию тяжелого заболевания – диабета.
Щитовидная железа находится в переднем средостении и состоит из двух или трех неравных долек. Структурно-функциональной единицей этого органа являются фолликулы, представляющие собой округлые полости различной величины. Между фолликулами находится соединительная ткань. Стенка фолликула щитовидной железы образована однослойным эпителием, расположенным на базальной пластинке. Полость фолликула заполнена коллоидом – полупрозрачным веществом, состоящим из белка тироглобулина (660 кД). Тироглобулин синтезируется и выделяется клетками фолликулярного эпителия – тироцитами. Одновременно тироциты поглощают тироглобулин и, расщепляя его, секретируют в кровь два гормона – тироксин (Т4) и трийодтиронин(Т3). Таким образом, сначала тироциты создают запас высокомолекулярного тироглобулина, а затем используют его для образования низкомолекулярных гормонов.
Активной формой гормона щитовидной железы является Т3. Поступая в ядра всех клеток организма, он регулирует интенсивность транскрипции генов и тем самым уровень обмена веществ в организме. Т4 служит в качестве неактивной формы для быстрой конвертации в Т3. Оба гормона содержат атомы йода, который является лимитирующим фактором для образования активной формы. Поэтому при недостатке йода в пище уровень обмена веществ снижается, а щитовидная железа компенсаторно увеличивается. Нарушения синтеза и выделения гормонов этой железой приводит к ряду тяжелых нарушений обмена веществ, замедленному развитию и умственной отсталости.
В состав щитовидной железы входят также группы клеток паращитовидной (околощитовидной) железы. Они секретируют в кровь зависимый от витамина D паратгормон, который способствует повышению концентрации кальция в крови. Его антагонистом является кальцитонин, который выделяется С-клетками фолликулярного эпителия щитовидной железы.
Дата добавления: 2014-02-04; просмотров: 6569; Опубликованный материал нарушает авторские права? | Защита персональных данных | ЗАКАЗАТЬ РАБОТУ
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: При сдаче лабораторной работы, студент делает вид, что все знает; преподаватель делает вид, что верит ему. 9479 — | 7337 — или читать все…
Читайте также:
Источник
Материал взят с сайта www.hystology.ru
Способность клеток организма интенсивно синтезировать активные вещества (секрет, гормон), необходимые для осуществления функции других органов, характерна для эпителиальной ткани. Эпителии, вырабатывающие секреты, называются железистыми, а его клетки — секреторными клетками, или секреторными гландулоцитами. Из секреторных клеток построены железы, которые могут быть оформлены в виде самостоятельного органа или являться только его частью.
Различают эндокринные (endo — внутри, krio — отделяю) и экзокринные (ехо — снаружи) железы. Экзокринные железы состоят из двух частей: концевой (секретирующей) части и выводных протоков, по которым секрет поступает на поверхность организма или в полость внутреннего органа. Выводные протоки обычно не принимают участие в образовании секрета.
Эндокринные железы лишены выводных протоков. Их активные вещества (гормоны) поступают в кровь, в связи с чем функцию выводных протоков выполняют капилляры, с которыми железистые клетки очень тесно связаны. Подробно функциональная морфология желез внутренней секреции будет рассмотрена в главе 8.
Экзокринные железы разнообразны по строению и функции. Они могут быть одноклеточными и многоклеточными. Примером одноклеточных желез служат бокаловидные клетки, встречающиеся в простом столбчатом каемчатом и псевдомногослойном реснитчатом эпителиях. Несекретирующая бокаловидная клетка цилиндрической формы и сходна с несекреторными эпителиоцитами. Секрет (муцин) накапливается в апикальной зоне, а ядро и органеллы смещаются к базальной части клетки. Смещенное ядро приобретает форму полулуния, а клетка — бокала. Затем секрет изливается из клетки, а она вновь приобретает столбчатую форму.
Экзокринные многоклеточные железы могут быть однослойными и многослойными, что обусловлено генетически. Если железа развивается из многослойного эпителия (потовая, сальная, молочная, слюнные железы), то и железа многослойна; если из однослойного (железы дна желудка, матки, поджелудочная железа), то они однослойны.
Характер ветвления выводных протоков экзокринных желез различен, поэтому они подразделяются на простые и сложные. У простых желез неветвящийся выводной проток, у сложных — ветвящийся.
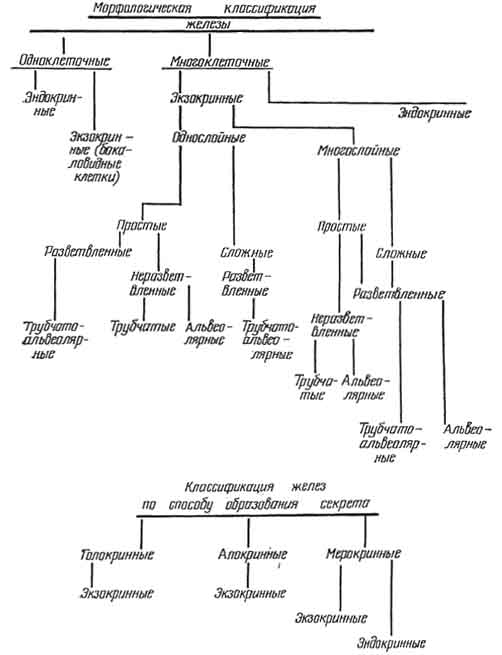
Рис. 92. Классификация желез.
Концевые отделы у простых желез разветвляются и не разветвляются, у сложных — разветвляются. В связи с этим у них и соответствующие названия: разветвленная железа и неразветвленная железа.
По форме концевых отделов экзокринные железы классифицируют на альвеолярные, трубчатые, трубчато-альвеолярные. У альвеолярной железы клетки концевых отделов формируют пузырьки или мешочки, у трубчатых — образуют вид трубочки. Форма концевой части трубчато-альвеолярной железы занимает промежуточное положение между мешочком и трубочкой (рис. 92, 93).
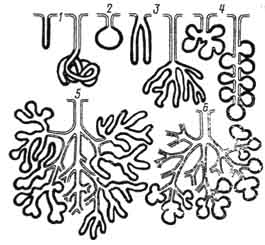
Рис. 93. Схематическое изображение простых и сложных экзокринных желез:
1 — простые трубчатые железы с неразветвленными концевыми отделами; 2 — простая альвеолярная железа с неразветвленным концевым отделом; 3 — простые трубчатые железы с разветвленными концевыми отделами; 4 — простые альвеолярные железы с разветвленными концевыми отделами; 5 — сложная альвеолярно-трубчатая железа с разветвленным концевым отделом; 6 — сложная альвеолярная железа с разветвленными концевыми отделами. Секреторные отделы изображены черными линиями, выводные протоки — светлыми.
Клетки концевого отдела именуются гландулоцитами. Процесс синтеза секрета начинается с момента поглощения гландулоцитами из крови и лимфы исходных компонентов секрета. При активном участии органелл, синтезирующих секрет белкового или углеводного характера, в гландулоцитах образуются секреторные гранулы. Они накапливаются в апикальной части клетки, а затем путем обратного пиноцитоза выделяются в полость концевого отдела. Завершающий этап секреторного цикла — восстановление клеточных структур, если в процессе секреции они разрушились.
Строение клеток концевой части экзокринных желез обусловлено составом выделяемого секрета и способом его образования.
По способу образования секрета железы делят на голокринные, апокринные, мерокринные (эккринные). При голокринной секреции (holos — целый) железистый метаморфоз гландулоцитов начинается с периферии концевого отдела и протекает в направлении выводного протока. Примером голокринной секреции является сальная железа. Стволовые клетки с базофильной цитоплазмой и округлым ядром расположены на периферии концевой части. Они интенсивно делятся митозом, поэтому мелкие по размеру. Перемещаясь к центру железы, секреторные клетки увеличиваются, так как в их цитоплазме постепенно накапливаются капельки кожного жира. Чем больше откладывается в цитоплазме жировых капель, тем интенсивнее протекает процесс деструкции органелл. Он завершается полным разрушением клетки. Плазмолемма разрывается, а содержимое гландулоцита поступает в просвет выводного протока.
При апокринной секреции (аро — от, сверху) разрушается апикальная часть секреторной клетки, являясь затем составной частью ее секрета. Данный тип секреции совершается в потовой или молочной железах.
При мерокринной секреции клетка не разрушается. Такой способ образования секрета типичен для многих желез организма: железы желудка, слюнные железы, поджелудочная железа, эндокринные железы (рис. 94).
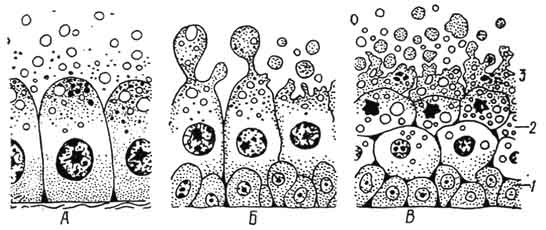
Рис. 94. Различные тппы секреции:
А — мерокринный; F — апокринный; В — голокринный; 1 — малодифференцированные клетки; 2 — перерождающиеся клетки; 3 — разрушающиеся клетки.
Таким образом, железистый эпителий так же, как и покровный, развивается из всех трех зародышевых листков (эктодермы, мезодермы, энтодермы), расположен на соединительной ткани, лишен кровеносных сосудов, поэтому питание осуществляется диффузионным способом. Клеткам свойственна полярная дифференцировка: в апикальном полюсе локализуется секрет, в базальном полюсе — ядро и органеллы.
Регенерация. Покровные эпителии занимают пограничное положение. Они часто повреждаются, поэтому характеризуются высокой регенерационной способностью. Регенерация осуществляется главным образом митомическим и очень редко амитотическим способом. Клетки эпителиального пласта быстро изнашиваются, стареют и гибнут. Их восстановление называется физиологической регенерацией.
Восстановление эпителиальных клеток, утраченных по причине травмы и другой патологии, называется репаративной регенерацией.
В однослойных эпителиях регенерационной способностью обладают или все клетки эпителиального пласта, или, если эпптелиоциты высокодифференцированны, то за счет зонально лежащих своих стволовых клеток.
В многослойных эпителиях стволовые клетки находятся на базальной мембране, поэтому лежат в глубине эпителиального пласта.
В железистом эпителии характер регенерации обусловлен способом образования секрета. При голокринной секреции стволовые клетки находятся снаружи железы на базальной мембране. Делясь и дифференцируясь, стволовые клетки преобразуются в железистые.
В мерокринных и апокринных железах восстановление эпителиоцитов протекает главным образом путем внутриклеточной регенерации.
Отзывов (0)
Добавить отзыв
Источник

