Действие фосфолипаз поджелудочной железы
В
процессах пищеварения все омыляемые
липиды (жиры, фосфолипиды, гликолипиды,
стериды) подвергаются гидролизу на
составные части, уже названные ранее,
стерины же химическим изменениям не
подвергаются. При изучении этого
материала следует обратить внимание
на отличия пищеварения липидов от
соответствующих процессов для углеводов
и белков: особую роль желчных кислот в
распаде липидов и транспорте продуктов
пищеварения. В составе липидов пищи
преобладают триглицериды. Фосфолипидов,
стреинов и других липидов потребляется
значительно меньше.
Большая
часть поступающих с пищей триглицеридов
расщепляется до моноглицеридов и жирных
кислот в тонком кишечнике. Гидролиз
жиров происходит под влиянием липаз
сока поджелудочной железы и слизистой
оболочки тонкого кишечника. Соли желчных
кислот и фосфолипиды, проникающие из
печени в просвет тонкого кишечника в
составе желчи, способствуют образованию
устойчивых эмульсий. В результате
эмульгирования резко увеличивается
площадь соприкосновения образовавшихся
мельчайших капелек жира с водным
раствором липазы, и этим самым увеличивается
липолитическое действие фермента. Соли
желчных кислот стимулируют процесс
расщепления жиров не только участвуя
в их эмульгировании, но и активируя
липазу. Расщепление стероидов происходит
в кишечнике при участии фермента
холинэстеразы, выделяющегося с соком
поджелудочной железы. В результате
гидролиза стероидов образуются жирные
кислоты и холестерин. Фосфолипиды
расщепляются полностью или частично
под действием гидролитических ферментов
— специфических фосфолипаз. Продуктом
полного гидролиза фосфолипидов являются:
глицерин, высшие жирные кислоты, фосфорная
кислота и азотистые основания.
Всасыванию
продуктов переваривания жиров предшествует
образование мицелл — надмолекулярных
образований или ассоциатов. Мицеллы
содержат в качестве основного компонента
соли желчных кислот, в которых растворены
жирные кислоты, моноглицериды, холестерин
и т.п.
В
клетках кишечной стенки из продуктов
пищеварения, а в клетках печени, жировой
ткани и других органов из предшественников,
возникших в обмене углеводов и белков,
происходит построение молекул
специфических липидов тела человека —
ресинтез триглицеридов и фосфолипидов.
Однако их жирнокислотный состав по
сравнению с жирами пищи изменен: в
триглицеридах, синтезируемых в слизистой
оболочке кишечника содержатся арахидоновая
и линоленовая кислоты даже в том случае,
если они отсутствуют в пище.
Фосфолипазы
— ферменты класса гидролаз, катализирующие
катаболизм глицерофосфолипидов.
Различают фосфолипазы секреторные,
входящие в состав панкреатического
сока, и клеточные фосфолипазы. Клеточные
фосфолипазы А1,
A2,
D, С различаются по специфично- к
отщепляемой группе. Все фосфолипазы —
кальцийзависимые ферменты.
Фосфолипаза
С —
фермент, гидролизующий фосфоэфирную
связь в глицерофосфолипидах. В клетках
человека идентифицировано 10 изоформ
фосфолипазы С, разлетающихся по
молекулярной массе, локализации, способу
регуляции, субстратной специфичности.
В структуре всех изоформ фосфолипазы
С отсутствуют гидрофобные домены,
которые могли бы обеспечить их
взаимодействие с мембраной. Однако
некоторые формы фосфолипазы С связаны
с мембраной с помощью гидрофобного
«якоря» — ацильного остатка
миристиновой кислоты или за счёт
взаимодействия с поверхностью бислоя.
Каталитическая активность всех изоформ
фосфолипазы С зависит от ионов кальция.
Большинство фосфолипаз С специфично в
отношении фосфатидилинозитолов и
практически не гидролизует другие типы
фосфолипидов. Активный фермент может
гидролизовать до 50% от общего количества
фосфатидилинозитолов клеточной мембраны.
При гидролизе фосфатидилинозитол-4,5-бисфосфата
(ФИФ2)
образуются продукты диацилглицерол
(ДАТ) и инозитол-1,4,5-трифосфат (ИФ3),
служащие вторичными посредниками в
трансмембранной передаче сигнала по
инозитолфосфатному пути.
В
переваривании глицерофосфолипидов
участвуют несколько ферментов,
синтезирующихся в поджелудочной железе.
Фосфолипаза
А2 гидролизует
сложноэфирную связь у второго атома
углерода глицерола, превращая
глицерофосфолипиды в соответствующие
лизофосфолипиды. Фосфолипаза
A2 секретируется
в кишечник в виде профермента и
активируется уже в полости кишечника
путём частичного протеолиза. Для
проявления активности фосфолипазы
A2 необходимы
ионы кальция.
Жирная
кислота в положении 1 отщепляется под
действием лизофосфолипазы, а
глицерофосфохолин гидролизуется далее
до глицерола, холина и фосфорной кислоты,
которые всасываются. Лизофосфолипиды
— эффективные эмульгаторы жира, ускоряющие
его переваривание.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Панкреатическая
липаза выделяется в полость тонкой
кишки из поджелудочной железы вместе
с белком колипазой. Колипаза попадает
в полость кишечника в неактивном виде
и частичным протеолизом под действием
трипсина превращается в активную форму.
Колипаза своим гидрофобным доменом
связывается с поверхностью мицеллы
эмульгированного жира. Другая часть
молекулы способствует формированию
такой конформации панкреатической
липазы, при которой активный центр
фермента максимально приближен к своим
субстратам — молекулам жиров, поэтому
скорость реакции гидролиза жира резко
возрастает. Панкреатическая липаза
гидролизует жиры преимущественно в
положениях 1 и 3, поэтому основными
продуктами гидролиза являются свободные
жирные кислоты и 2-моноацилглицеролы
(β-моноацилглицеролы). Молекулы
2-моноацилглицеролов также обладают
детергентными свойствами и способствуют
эмульгированию жиров.
В
переваривании глицерофосфолипидовучаствуют
несколько ферментов, синтезирующихся
в поджелудочной железе. Фосфолипаза А2
гидролизует сложноэфирную связь у
второго атома углерода глицерола,
превращая глицерофосфолипиды в
соответствующие лизофосфолипиды.
ФосфолипазаA2 секретируется в кишечник
в виде профермента и активируется уже
в полости кишечника путём частичного
протеолиза. Для проявления активности
фосфолипазыA2 необходимы ионы кальция.
Жирная кислота в положении 1 отщепляется
под действием лизофосфолипазы, а
глицерофосфохолин гидролизуется далее
до глицерола, холина и фосфорной кислоты,
которые всасываются. Лизофосфолипиды
— эффективные эмульгаторы жира, ускоряющие
его переваривание.
Переваривание
эфиров холестерола
В
составе пищи холестерол находится в
основном в виде эфиров. Гидролиз эфиров
холестерола происходит под действием
холестеролэстеразы — фермента, который
также синтезируется в поджелудочной
железе и секретируется в кишечник.
Продукты гидролиза (холестерол и жирные
кислоты) всасываются в составе смешанных
мицелл. Следствие этого нарушение
переваривания и всасывания жиров у
детей более опасно, чем у взрослых.
Всасывание
продуктов гидролиза липидов в тонком
кишечнике.
Образование
смешанных мицелл.
Продукты
гидролиза липидов — жирные кислоты с
длинным углеводородным радикалом,
2-моноацилглицеролы, холестерол, а также
соли жёлчных кислот образуют в просвете
кишечника структуры, называемые
смешанными мицеллами. Смешанные мицеллы
построены таким образом, что гидрофобные
части молекул обращены внутрь мицеллы,
а гидрофильные — наружу, поэтому мицеллы
хорошо растворяются в водной фазе
содержимого тонкой кишки. Стабильность
мицелл обеспечивается в основном солями
жёлчных кислот. Мицеллы сближаются со
щёточной каймой клеток слизистой
оболочки тонкого кишечника, и липидные
компоненты мицелл диффундируют через
мембраны внутрь клеток. Вместе с
продуктами гидролиза липидов всасываются
жирорастворимые витамины A, D, Е, К и соли
жёлчных кислот. Наиболее активно соли
жёлчных кислот всасываются в подвздошной
кишке. Жёлчные кислоты далее попадают
через воротную вену в печень, из печени
вновь секретируются в жёлчный пузырь
и далее опять участвуют в эмульгировании
жиров. Этот путь жёлчных кислот называют
«энтерогепатическая циркуляция».
Каждая молекула жёлчных кислот за сутки
проходит 5- 8 циклов, и около 5% жёлчных
кислот выделяется с фекалиями. Всасывание
жирных кислот со средней длиной цепи,
образующихся, например, при переваривании
липидов молока, происходит без участия
смешанных мицелл. Эти жирные кислоты
из клеток слизистой оболочки тонкого
кишечника попадают в кровь, связываются
с белком альбумином и транспортируются
в печень.
Соседние файлы в предмете Биохимия
- #
- #
- #
- #
- #
- #
Источник
Содержание
Введение
1. Фосфолипазы
1.1 Классификация. Свойства
1.2 Система фосфолипаза С — инозитол-3-фосфат
2. Фосфолипазы А2
2.1 Общие сведения (реакция, открытие, строение)
2.2 Классификация и свойства
2.2.1 Цитозольные ФЛА2
2.2.2 Секреторные ФЛА2
2.2.3 Кальцийнезависимая ФЛА2
2.3 Субстратная специфичность
2.4 Ингибиторы ФЛА2
2.4.1 Неконкурентное ингибирование
2.4.2 Конкурентное ингибирование
2.5 Значение для организма при нарушении активности
2.6 Использование ФЛА2 в медицине
2.7 Биологическая роль ФЛА2
Список литературы
Введение
Фосфолипазы (англ. phospholipase) ферменты класса гидролаз, катализирующие гидролиз фосфоглицеридов.. В зависимости от положения гидролизуемой связи в фосфолипиде различают 4 основных класса фосфолипаз: A, B, C и D.

Лизофосфолипиды расщепляются под действием фосфолипаз L (существование позиционно специфичных фосфолипаз L1 и L2 не доказано). Фосфолипазы В — устаревшее назв. препаратов, обладающих активностью по типу фосфолипаз А и L.

X — остаток холина, серина, миоинозита и др.; для фосфолипаз L1 R2=C(O)R4, R3=H; для фосфолипаз L2 R2=H, R3=C(O)R4
Каждое из семейств фосфолипаз неоднородно и включает ферменты, значительно отличающиеся по молекулярным массам, субъединичному составу и другим свойствам. Все фосфолипазы наиболее активно катализируют гидролиз на поверхности раздела фаз фосфолипид — вода; медленно гидролизуют водорастворимые субстраты.
Фосфолипаза A1
— (КФ 3.1.1.32, англ. phospholipase A1) отщепляет SN-1 ацильную цепь.
Фосфолипаза A2
— (КФ 3.1.1.4, англ. phospholipase A2) отщепляет SN-2 ацильную цепь.
Фосфолипаза B
-(лизофосфолипаза, англ. phospholipase B) отщепляет обе SN-1 и SN-2 ацильные цепи. Фосфолипаза, обладающая активностями как фосфолипазы А1 так и А2, то есть способной гидролизовать ацильную цепь фосфолипида в sn-1 и sn-2 положениях.
Фосфолипаза C
— (КФ 3.1.4.3, англ. phospholipase C) гидролизует связь между глицериновым остатком фосфолипида и полярной фосфатной группой, при этом образуются диацилглицерин и фосфат-содержащая полярная группа.
Фосфолипаза D
— (КФ 3.1.4.4, англ. phospholipase D) гидролизует связь между фоасфатной группой и спиртовой группой, при этом высвобождаются фосфатидная кислота и спирт. Существует 2 изоформы этой фосфолипазы D1 и D2.
Фосфолипазы играют важную роль в обмене липидов в живых организмах. Их используют для определения структуры фосфоглицеридов и места их локализации в мембранах.
1. Фосфолипазы.
1.1 Классификация. Свойства
Фактически различают несколько фосфолипаз группы А, они являются составной частью многих тканей и секретов живых организмов.
Фосфолипазы A1
в большинстве своем — внутриклеточные ферменты, часто мембраносвязанные, не нуждаются в коферменте. Их молекулярные массы варьируют в пределах 15-90 тыс.; оптимальная каталитическая активность проявляется при рН 4,0 (для лизосомальных ферментов) или 8,0-9,5 (для ферментов микросом, плазматических мембран и цитозоля); широко распространены в животных тканях (печень, сердце, мозг) и в микроорганизмах (Bacillus subtilis, В. megateiium, Mycobacter phlei, Escherichia coli).
Фосфолипазы А1 отщепляют ацильную цепь фосфолипида в sn-1 положении. При действии фосфолипазы А1 на фосфолипид образуется 1-лизофосфолипид и жирная кислота. Фосфолипаза является активным компонентом змеиного яда гемолитического действия.
Фосфолипазы A2
— наиболее изученные представители фосфолипаз. Известны 3 группы фосфолипаз A2: 1) ферменты ядов змей, рептилий и насекомых, существующие в виде большого количества изоформ; 2) ферменты поджелудочной железы млекопитающих, продуцирующиеся в организме в виде зимогенов (предшественников с большей молекулярной массой) и активирующиеся трипсином; 3) внутриклеточные ферменты из крови и тканей животных, среди которых имеются как растворимые, так и мембраносвязанные.
Фосфолипазы A2 первых двух подгрупп являются водорастворимыми ферментами с молеклярной масссой 11-19 тыс. (некоторые активны в виде димеров), обладают высокой стабильностью благодаря большому числу (6-7) дисульфидных связей. Оптимальная каталитическая активность при рН 7,5-9,0; рI от 4,0 до 10,5; кофермент — Ca2+. Для множества представителей этих подгрупп фосфолипаз известны первичная и пространственная структур. В активном центре обнаружены остатки гистидина и аспарагиновой кислоты. Свойства внутриклеточных фосфолипаз A2 (третья подгруппа) зависят от субклеточной локализации фермента. Их молекулярная масса 12-75 тыс.; оптимальная каталитическая активность при рН 4,2-9,0. Некоторые ферменты этой подгруппы не содержат коферментов.
Фосфолипазы В
выделены из растений, микроорганизмов, яда пчел, тканей млекопитающих. Ферменты этой группы крайне неспецифичны, катализируют гидролиз различных сложноэфирных связей, обладают литическим (разрушающим) действием по отношению к биологически мембранам (что обусловливает их токсичность). Молекулярная масса фосфолипаз В 15-65 тыс., они менее стабильны, чем фосфолипазы А; их оптимальная каталитическая активность проявляется при рН от 4,5 (лизосомальный фермент) до 10,0 (ферменты ядов). Фосфолипазы не имеют коферментов, не ингибируются этилендиаминтетрауксусной кислотой. Некоторые фосфолипазы В ингибируются диизопропилфторфосфатом и п-хлормеркурбензойной кислотой. Универсальные ингибиторы для всех фосфолипаз В — ПАВ.
Фосфолипаза В способна гидролизовать ацильную цепь фосфолипида в sn-1 и sn-2 положениях Как правило фосфолипаза действует на лизолецитин (лизофосфатидилхолин), образующийся в результате действия фосфолипазы А1 на лецитин (фосфатидилхолин).
Фосфолипазы С
обнаружены у бактерий Clostridium, Bacillus и Pseudomonas, а также в клетках млекопитающих (печень, мозг, поджелудочная железа). Для некоторых из них характерна строгая специфичность по отношению к спиртовой группе молекулы субстрата, например к остатку холина (фосфолипазы Cx) и миоинозита (фосфолипазы Си). Молекулярная масса фосфолипаз С от 23 до 51 тыс. Ионы Zn2+ являются для них коферментом и стабилизатором. Оптимальная каталитическая активность при рН около 7 для фосфолипаз Cx, и при рН < 7 для фосфолипаз Си.
Фосфолипаза С, гидролизующая фосфодиэфирную связь между глицериновым остатком фосфолипида и полярной фосфатной группой, относится к фосфодиэстеразам также как и фосфолипаза D. Фосфолипаза С является ключевым ферментом метаболизма фосфатидилинозитола и липидных сигнальных путей.
Фосфолипаза С активируется Gαq или Gβγ субъединицами G-белка. Таким образом, она является частью G-белоксвязанного рецептора (англ. G protein-coupled receptor) и соответствующего сигнального пути или частью трансмембранного рецептора с внутренней или ассоциированной тирозинкиназной активностью.
Фосфолипаза С гидролизует фосфатидилинозитол (PIP2) на два вторичных медиатора инозитолтрифосфат (IP3) и диацилглицерин (DAG). Эти медиаторы становятся вовлечены в последующие этапы сигнальных путей. В частности, они модулируют кальциевые каналы эндоплазматического ретикулума и протеинкиназу С, соответственно.
Фосфолипаза D
относится к группе важных ферментов, которые в живых системах выполняют разнообразные функции от усвоения питательных веществ до синтеза биологически активных соединений. Обнаружены в растениях (овощи, водоросли), микроорганизмах и в тканях животных. Их молекулярная масса 90-116 тыс. Оптимальная каталитическая активность при рН 4,7-8,0. Катионные ПАВ ингибируют фосфолипазы D, анионные — активируют.
Фосфолипаза D проявляет прежде всего гидролитическую активность, в результате которой происходит расщепление сложноэфирной связи между остатком фосфатидной кислоты и спирта в молекулах фосфолипидов (ФЛ). При этом последний замещается на водород, но возможен перенос остатка фосфатидной кислоты на самые разные гидроксилсодержащие акцепторы, что представляет большой интерес для биотехнологии, так как трансфосфатидилирующая активность фосфолипазы может быть использована для синтеза разнообразных лекарственных препаратов.
Фосфолипаза D специфически расщепляет фосфатидилхолин на фосфатидную кислоту и холин, высвобождая последний в цитоплазму.



фосфатидилхолин фосфатидная кислота холин
1.2 Система фосфолипаза С — инозитол-3-фосфат
Активация мембранной гуанилатциклазы происходит не под непосредственным влиянием гормон-рецепторного комплекса, а опосредованно через ионизированный кальций и ок-сидантные системы мембран. Определяющая эффекты ацетилхолина стимуляция активности гуанилатциклазы также осуществляется опосредованно через Са2+. Через активацию гуанилатциклазы реализует эффект и на-трийуретический гормон предсердий — атриопептид. Путем активации пе-рекисного окисления стимулирует гуанилатциклазу гормон эндотелия сосудистой стенки оксид азота — расслабляющий эндотелиальный фактор. Под влиянием гуанилатциклазы из ГТФ синтезируется цГМФ, активирующий цГМФ-зависимые протеинкиназы, которые уменьшают скорость фосфорилирования легких цепей миозина в гладких мышцах стенок сосудов, приводя к их расслаблению.
В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны. Примерами могут служить стимуляция сокращений сердца под влиянием цАМФ и торможение их цГМФ, стимуляция сокращения гладких мышц кишечника цГМФ и подавление цАМФ. цГМФ обеспечивает гиперполяризацию рецепторов сетчатки глаза под влиянием фотонов света. Ферментативный гидролиз цГМФ, а следовательно, и прекращение гормонального эффекта, осуществляется с помощью специфической фосфодиэстеразы.

Источник
Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 | |
Источник: biochemistry.ru
61
Меньшая часть (40%) моноацилглицеролов подвергается дальнейшему гидролизу до глицерола.
Для остальной части процесс ферментативного гидролиза завершается на этапе образования 2- моноацилглицеролов.
В расщеплении МАГ участвует также кишечная липаза, но активность этого фермента невысока.
62
31

Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
В соке поджелудочной железы присутствуют и другие ферменты, способные расщеплять липиды. В частности, эстеразы катализируют преимущественно гидролиз эфиров жирных кислот с короткой цепью.
В поджелудочной железе синтезируется профосфолипаза А2.
63
64
32
Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
Фермент приобретает активность только после воздействия в просвете кишечника трипсина, приводящего к отщеплению от него гептапептида.
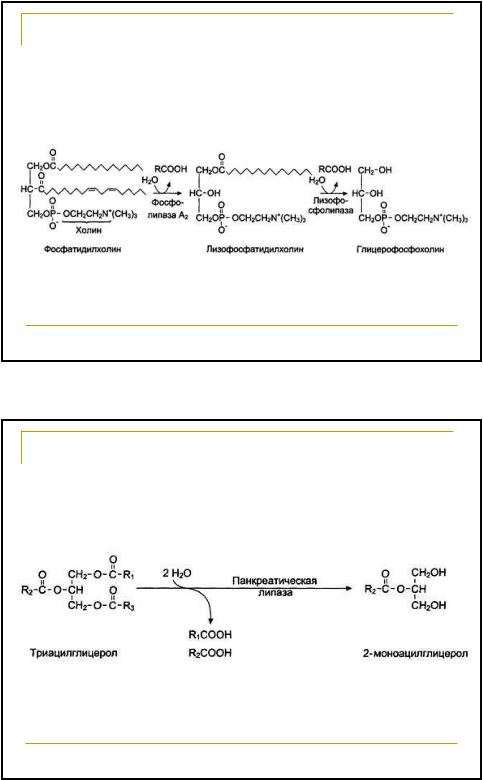
Фосфолипаза А2 катализирует отщепление молекулы жирной кислоты от фосфатидилхолина с образованием лизофосфатидилхолина
65 | |||
Расщепление ФЛ пищи происходит под | |||
действием фосфолипаз | |||
панкреатического сока | |||
Фосфолипаза А2 | Фосфолипаза А1 | ||
О | СН2-О-СО-R1 | ||
R2-C-O-CH | О | ||
CH2-O-Р-О-Х | |||
Фосфолипаза С | О- | Фосфолипаза Д | |
66 | |||
33
Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
Образующиеся при этом продукты называются
лизофосфолипидами. Так, при гидролизе фосфатидилхолина образуются
лизофосфатидилхолин и лизофосфатидилэтаноламин. Они токсичны и вызывают разрушение мембран клеток.
Именно поэтому яд змей (кобра, гюрза) содержит высокоактивную фосфолипазу А2, вызывающую гемолиз эритроцитов.
67
В расщеплении жиров принимает участие и кишечная липаза, однако активность ее мала, и она гидролизует расщепление
только моноглицеридов (МГ).
Т.о., продуктами гидролиза ТГ являются
ЖК, МГ и глицерин (Гн)
69
34

Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
Всасывание:
Тонко эмульгированные жиры (размер капель эмульсии менее
0,5 мкм) частично всасываются без предварительного гидролиза.
ЖК с короткой углеродной цепью
(менее С12) и Гн хорошо растворимы в воде, свободно всасываются стенкой кишечника и далее поступают в v. portae, затем в печень.
71
ЖК с длинной цепью и МАГ всасываются с помощью:
желчных кислот,
ФЛ (фосфолипидов),
ХС (холестерина).
Из них образуются мицеллы.
72
35

Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
Структура этих частиц такова, что их гидрофобное ядро (ЖК, МГ) оказывается окруженным гидрофобной оболочкой из желчных кислот и ФЛ. Всасывание происходит путем мицеллярной диффузии или путем пиноцитоза.
73
Однако в кишечнике, при одновременном действии обеих фосфолипаз А1 иА2, токсическое действие снимается. Холестерин( ХС), поступающий с пищей( мясо, яйца) либо в свободном состоянии, либо в виде эфиров, всасывается только в присутствии желчных кислот.
Эфиры ХС при этом могут разрушаться с помощью холестеролэстеразы.
74
36

Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
Ресинтез липидов в стенке кишечника.
С пищей попадают разнообразные липиды, в том числе и чужеродные для организма.
В стенке кишечника происходит ресинтез специфичных для данного организма липидов.
75
Это обеспечивается тем, что в синтезе ТГ и ФЛ принимают участие как экзогенные, так и эндогенные ЖК.
Однако способность организма к ресинтезу липидов ограничена. Часть пищевого чужеродного жира все же откладывается в жировых депо.
76
37
Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
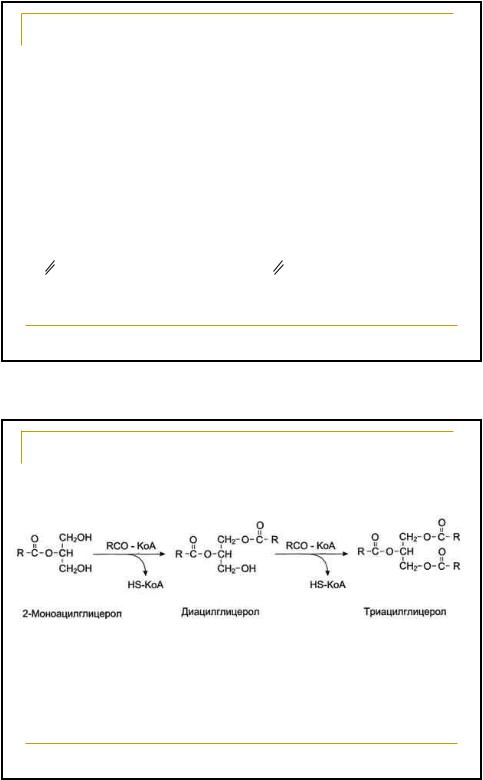
Механизм ресинтеза | |
Всосавшиеся ЖК активируются. | |
Активация их заключается в | |
присоединении остатка жирной | |
кислоты к коферменту А с | |
образованием ацил~КоА. | |
Это происходит в гладком | |
эндоплазматическом ретикулуме | |
O | O |
R-C- O- + АТФ + HS-KoA R-C ~ S-KoA + АМФ + ФФн | |
77 | |
Ресинтез жиров в клетках слизистой | |
оболочки тонкой кишки | |
78
38
Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 | ||||||
переваривание, | |||||||
всасывание | |||||||
Н2С-ОН | |||||||
глицерол | НС-ОН | ||||||
АТФ | Н2С-ОН | ||||||
глицеролкиназа | |||||||
АДФ | |||||||
Н2С-ОН | Н2С-ОН | ||||||
глицерол-3-Р НС-ОН | 2-ацилглицеролНС-О-СO-R2 | ||||||
R1-CO~SKoA | Н2С-О-P | Н2С-ОH | |||||
HSKoA | глицерол-3-P- | а | |||||
ацилтрансфераза | |||||||
ц | |||||||
Н2С-О-СО-R1 | |||||||
и | |||||||
лизофосфолипид НС-ОН | л | ||||||
т | |||||||
R2-CO~SKoA | Н2С-О-P | р | |||||
1-ацилглицерол-3-P- | а | ||||||
HSKoA | ацилтрансфераза | н | |||||
Н2С-О-СО-R1 | с | ||||||
фосфатидная | ф | ||||||
кислота | НС-О-СO-R2 | е | |||||
Н2С-О-P | р | ||||||
Н2О | а | ||||||
фосфатидатфосфатаза | HS-KoA | R1-CO~SKoA | з | ||||
Рн | |||||||
а | |||||||
Н2С-О-СО-R1 | |||||||
диацилглицерол | НС-О-СO-R2 | Н2С-О-СО-R1 | |||||
холинфосфотрансфераза | |||||||
Н2С-ОН | |||||||
НС-О-СO-R2 | |||||||
диацилглицерол- | |||||||
ацилтрансфераза | Н2С-О-P-О-СН2-СН2-N(СН3)3 | ||||||
Н2С-О-СО-R1 | ЦДФ-холин | ЦМФ | фосфатидилхолин | ||||
(этаноламин) | |||||||
триацилглицерол НС-О-СO-R2 | |||||||
Н2С-О-СО-R3 | 79 | ||||||
В клетках слизистой тонкого | |||||||
кишечника функционируют два пути ресинтеза ТГ.
Это связано с поступлением туда при всасывании большого количества- 2-МАГ
80
39

Кафедра биохимии ГомГМУ, 2012 | 16.11.2012 |
Исключением, имеющим клиническое значение, является тот факт, что ТГ, содержащие короткие, и среднецепочечные жирные кислоты (С6-С10), могут подвергаться всасыванию без предварительного расщепления ферментами.
81
Из клеток слизистой тонкого кишечника они сразу попадают в кровоток системы воротной вены.
Этот механизм принципиально отличается от механизма всасывания и попадания в кровь основной массы липидов пищи, в составе
которых содержатся жирные кислоты с количеством углеродных атомов более> 10.
82
40
Источник

