
Главный стимулятор клеток поджелудочной железы называется ацетилхолин
Секреция ПЖ находится под контролем нервных и гуморальных механизмов. Начальную секрецию ПЖ вызывают вид, запах пищи и другие раздражители (условнорефлекторные сигналы), а также жевание и глотание (безусловнорефлекторные сигналы). При этом нервные сигналы, формирующиеся в рецепторах полости рта и глотки, достигают продолговатого мозга, и затем эфферентные влияния по волокнам блуждающего нерва поступают к железе и вызывают её секрецию.
У человека с фистулой ГПП выделение панкреатического сока начинается через 2—3 мин после того, как он увидел пищу или услышат о ней. Это пример условнорефлекторного пути возбуждения панкреатической секреции. Симпатические волокна, иннервирующие ПЖ, тормозят её секреторную активность и в то же время изменяют реактивность железы по отношению к другим воздействиям, усиливая синтез органических веществ.
Торможение панкреатической секреции происходит при раздражении многих чувствительных нервов, при болевых реакциях, во время сна, при напряжённой физической и умственной работе.
Для стимуляции панкреатической секреции прямые нервные влияния имеют меньшее значение, нежели гуморальные. Ведущее значение в гуморальной регуляции секреции ПЖ принадлежит желудочно-кишечным гормонам.
Большинство регуляторов секреции ферментов ПЖ действуют на рецепторы мембраны ацинарных клеток, расположенные на их базолатеральной поверхности. Выделяют рецепторы ХК, бомбезина, ацетилхолина, субстанции Р, ВИП, секретина.
Стимуляторы панкреатической секреции. ВИП и секретин стимулируют панкреатическую секрецию, активируя аденилатциклазу. Как и в других типах клеток, аденилатциклаза способствует образованию циклического аденозинмонофосфата, в результате чего протеинкиназа А, усиливающая секрецию панкреатического сока, богатого бикарбонатами, переходит в активную форму. Другие стимуляторы поджелудочной секреции (ХК, ацетилхолин, гастрин-рилизинг пептид, субстанция Р) действуют на специфические рецепторы, во внутриклеточной передаче сигнала от которых задействованы альтернативные вторичные мессенджеры.
Эти вещества повышают внутриклеточное содержание циклического гуанозинмонофосфата, что приводит к увеличению внутриклеточного содержания инозитолтрифосфата, диацилглицерола, арахидоновой кислоты и кальция. Эти промежуточные вещества-посредники активируют различные протеинкиназы, в результате этого повышается секреция ферментов. Данные, полученные в опытах на животных, свидетельствуют, что действие комбинации агонистов на различные мембранные рецепторы может вызывать синергический, но не суммарный (аддитивный) эффект. Например, ХК увеличивает секрецию бикарбонатов, стимулированную секретином, но секретин не повышает секреторный ответ на действие ХК.
Ингибиторы секреции поджелудочной железы. Различные вещества, ответственные за подавление панкреатической секреции, действуют по принципу отрицательной обратной связи во время и после приёма пищи.
Панкреатический полипептид (ПП) представляет собой пептидный гормон, образующийся в островках Лангерганса и подавляющий панкреатическую секрецию воды, бикарбонатов и ферментов. Концентрация ПП в плазме крови возрастает после мнимого кормления, после приёма пищи, после экспериментального закисления среды ДПК, а также при стимуляции блуждающего нерва, при действии ХК, секретина, ВИП. ПП может выступать как антагонист мускариновых рецепторов и способен ингибировать выделение ацетилхолина из постганглионарных нейронов ПЖ; его конечный эффект проявляется на уровне ацинарных клеток.
Пептид YY высвобождается в дистальной части подвздошной кишки и и толстой кишке в ответ на поступление пищи смешанного характера, по жиры, находящиеся в просвете кишки, в большей степени способны стимулировать его секрецию. Пептид YY уменьшает чувствительность ПЖ к действию секретина и ХК, возможно, за счёт уменьшения секреции ацетилхолина и норадреналина и ингибирования выделения ХК слизистой оболочкой ДПК.
Соматостатин ингибирует секрецию секретина слизистой оболочкой ДПК, а также уменьшает чувствительность к секретину рецепторных полей. Единственный эффект соматостатина — снижение секреции ферментов и бикарбонатов ПЖ. Соматостатин синтезируют клетки слизистой оболочки желудка и кишечника, а также D-клетки островков Лангерганса. Его активность составляет только около 25% от активности гипоталамического соматостатина. Но только соматостатин, продуцируемый слизистой оболочкой тонкой кишки, оказывает угнетающее действие на секрецию ПЖ.
Выделение соматостатина происходит при участии автономной нервной системы в ответ на поступление жиров и аминокислот с пищей.
Соматостатин блокирует панкреатическую секрецию несколькими способами. Во-первых, он действует путём угнетения продукции стимулирующих пептидов (ХК). Во-вторых, посредством угнетающего воздействия на ЦНС и регулируя работу интрапанкреатических ганглиев, соматостатин ингибирует поступление ацетилхолина в пресинаптическую щель и, возможно, ингибирует выработку инсулина. Хотя рецепторы к соматостатину найдены также в ацинарных клетках, эти рецепторы скорее способствуют увеличению секреции, чем блокируют её.
Другие ингибиторы, представленные среди гормонов эндокринных клеток островков Лангерганса, включают панкреатический глюкагон и панкреастатин, а также нейропептиды: кальцнтонин-генерирующий пептид и энкефалины (табл. 1-3). Панкреатический глюкагон ингибирует секрецию ПЖ, стимулированную ХК, секретином или пищей; угнетает секрецию бикарбонатов, воды и ферментов. Панкреастатин ингибирует панкреатическую секрецию, тормозя высвобождение ацетилхолина эфферентными окончаниями блуждающего нерва. Кальцитонин-генерирующий пептид может проявлять свою активность через стимуляцию выделения соматостатина. Энкефалины и подобные им опиоиды снижают выделение секретина слизистой оболочкой ДПК и могут также ингибировать высвобождение ацетилхолина.
Таблица 1-3. Ингибиторные нейромодуляторы

Маев И.В., Кучерявый Ю.А.
Источник
Экзокринная
функция поджелудочной железы включает
в себя выработку пищеварительных
ферментов в ацинозных клетках, а также
секрецию воды и электролитов
(преимущественно бикарбонатов),
осуществляемую главным образом
эпителиальными клетками панкреатических
протоков. Жидкая часть секрета, содержащая
НС03″,
обеспечивает транспортировку
панкреатических ферментов в
двенадцатиперстную кишку и защищает
их от расщепления соляной кислотой
желудочного сока, обеспечивая при
рН 7—8 их оптимальную активность.
Снижение выработки панкреатических
бикарбонатов (например, при хроническом
панкреатите) приводит, кроме того,
к ацидификации (закислению) дуоденального
содержимого, способствуя тем самым
развитию язвенных поражений (так
называемых панкреатогенных язв). Общий
объем панкреатической секреции
достигает 2,5 л в сутки.В поджелудочной
железе вырабатывается большое число
различных ферментов, необходимых
для переваривания белков, жиров и
углеводов. Так, протеазы (трипсин,
химотрипсин, эластаза, карбоксипепти-
даза, калликреин) расщепляют белки на
олигопептиды; липолитические ферменты
(липаза, колипаза, фосфолипазаА2и
карбоксилэстергидрола-
за)
эмульгируют капельки жира в присутствий
желчных кислот и расщепляют
триглицериды и холестерин на моноглицериДЫ
и жирные кислоты; амилаза участвует в
переваривании крахмала и гликогена,
превращая эти углеводы в ди- и
олигосахариды.
Существуют
различные стимуляторы панкреатической
секреции. При этом вазоинтестинальный
пептид (ВИП) и секретин оказывают
стимулирующее влияние за счет
активации аденилатциклазы и последующего
образования цАМФ, который в свою очередь
активирует протеинкиназу А. Другие
стимуляторы (ацетилхолин, холецистокинин,
субстанция Р) воздействуют на
специфические рецепторы, в результате
чего увеличивается внутриклеточное
содержание промежуточных «мессенджеров»
(ино- зитол-трифосфата, Са++
и др.), также активирующих различные
протеинкиназы. Тормозящее влияние на
панкреатическую секрецию оказывают
панкреатический полипептид, соматостатин,
глюкагон, энкефа- лины и другие
нейропептиды,
Регуляция
внешнесекреторной функции поджелудочной
железы представлена на схеме 22.3. В
межпищеварительный период уровень
секреции панкреатических ферментов,
а также воды и электролитов очень
невелик, составляя соответственно 10 и
1—2 % от их максимально возможной
продукции. Пищеварительная секреция
поджелудочной железы включает в себя
несколько фаз.
Цефалическая
фаза инициируется представлениями о
пище, ее ви
—
Секреция | |||
Ам | Лннолитпческие | ||
Протеолптнческис
Кишечная
фаза
Секреция
бикарбонатов
Цефалическая
фаза Желудочная
фаза
-представления
о пище -растяжение
-пил
пищи желудка -запах пищи
-ВКУС
1ШИ1Г1
—
трипсин -оеамилаза — липаза
-химотрипсин -колипаза
-эластаза -фосфолипаза
А2
■
карбокашептидаза ‘
-карбоксилэстср-
-калликрени гид
рол а л а
Схема
22.3.
Регуляция внешнесекреторной функции
поджелудочной железы.
52
7
дом,
запахом и вкусом. Их возбуждающее
влияние реализуется через ветви
блуждающего нерва и приводит к
значительному увеличению секреции
бикарбонатов и умеренному повышению
выработки панкреатических.фер- ментов.
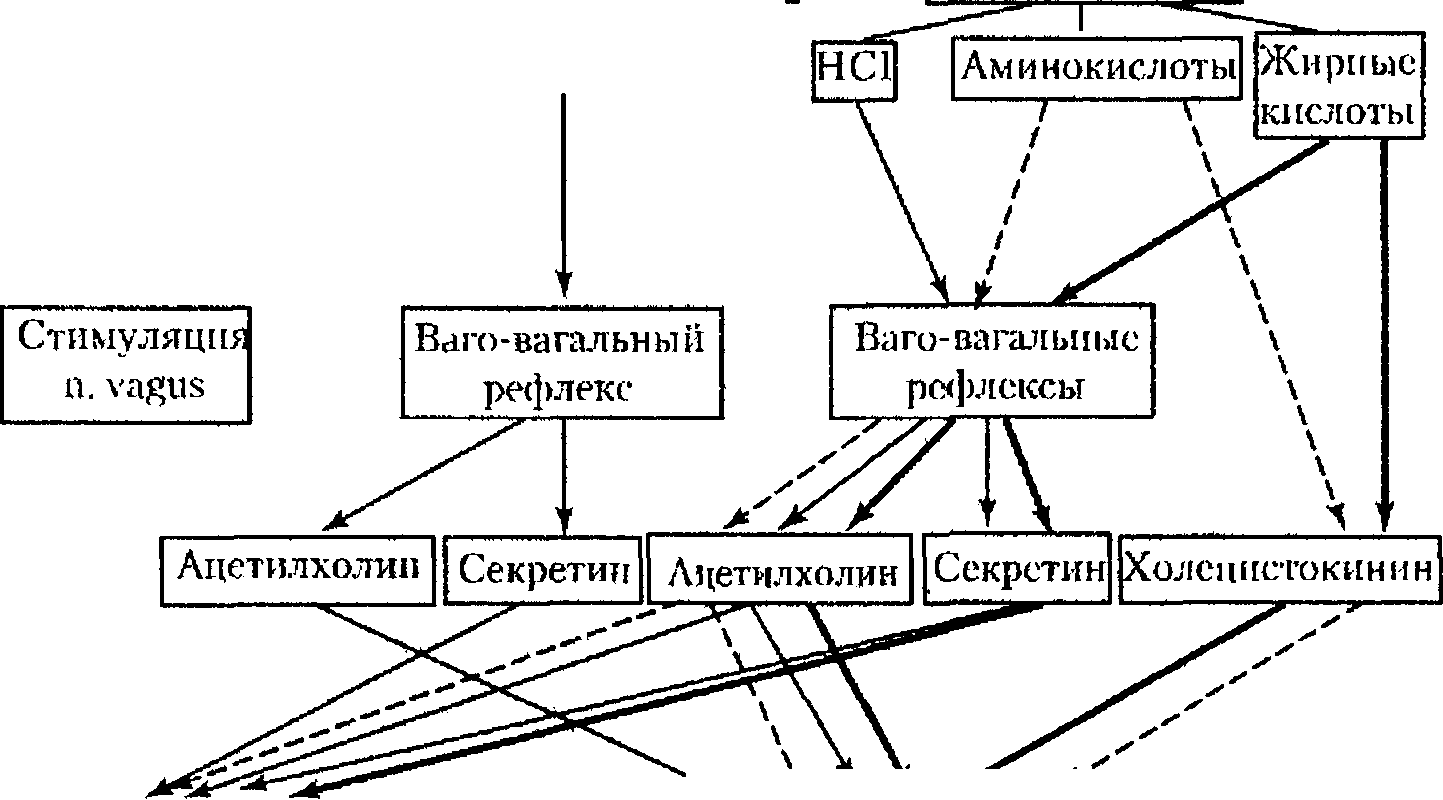
Желудочная
фаза включается — через ваго-вагальные
рефлексы — в ответ на растяжение пищей
фундального иантрального отделов
желудка, приводящее к освобождению
ацетилхолина и секретина и последующему
значительному увеличению продукции
панкреатических ферментов и очень
умеренному повышению секреции
бикарбонатов.
Интестинальная
(тонкокишечная) фаза панкреатической
секреции наиболее значима. В физиологических
условиях она сопровождается повышением
кровотока в поджелудочной железе и
выработкой панкреатического секрета
с очень высоким содержанием ферментов.
Выраженное стимулирующее действие на
секрецию поджелудочной железы (в
большей степени — панкреатических
ферментов, в меньшей — бикарбонатов)
оказывают продукты расщепления жиров,
в частности, жирные кислоты. Этот эффект
опосредуется через высвобождение
секретина, ацетилхолина и
холецистокинина. Соляная кислота,
попадая в двенадцатиперстную кишку,
приводит к ацидификации дуоденального
содержимого, вследствие чего
включается ваго-вагальный
энтеро-панкреатический рефлекс с
последующим высвобождением ацетилхолина
и секретина, способствующим выраженному
повышению секреции бикарбонатов и
умеренному увеличению выработки
панкреатических ферментов. Наконец,
пептиды и аминокислоты, также вызывая
ваго-вагальный рефлекс и при их приеме
в большом количестве повышая освобождение
холецистокинина, приводят к
значительному увеличению выработки
панкреатических ферментов и очень
умеренному возрастанию секреции
панкреатических бикарбонатов.
Нарушение
регуляции различных фаз панкреатической
секреции (например, усиленная стимуляция
внешнесекреторной функции поджелудочной
железы ацетилхолином, соляйОй кислотой,
холецистокинином и панкреозимином)
может приводить к развитию повреждения
паренхимы поджелудочной железы.
Это, в частности, находит свое отражение
при воспроизведении различных
экспериментальных моделей острого
панкреатита у животных. Однако
конкретное значение указанных нарушений
в развитии заболеваний поджелудочной
железы у человека остается пока еще
недостаточно изученным.
Соседние файлы в предмете Патологическая физиология
- #
23.02.201516.36 Mб258Patofiziologia_Zayko.pdf
- #
- #
- #
- #
- #
- #
Источник
Нервная и гуморальная регуляция секреторной функции поджелудочной железы
Слабое сокоотделение натощак резко усиливается во время (через 2— 3 мин) и после приема пищи. В привычных условиях приема пищи панкреатическое сокоотделение начинается уже на вид, запах пищи и другие раздражители, сопутствующие приему пищи, что свидетельствует об условно-рефлекторном механизме секреции. При раздражении пищевыми веществами рецепторов слизистой оболочки ротовой полости, глотки и пищевода возникает безусловно-рефлекторное отделение панкреатического сока. Нервные импульсы от раздражаемых рецепторов по афферентным путям достигают бульбарного центра панкреатической секреции, где они переключаются на преганглионарные нейроны ядер блуждающего нерва, по эфферентным волокнам которого достигают постганглионарных нейронов. Их аксоны образуют синаптические окончания на базальных мембранах панкреацитов. Выделяющийся при возбуждении этих окончаний ацетилхолин активирует М-холинорецепторы постсинаптической мембраны. При этом освобождаются вторичные посредники (Са++ и ГЦ-цГМФ), которые и вызывают секреторную деятельность панкреацитов и эпителиальных клеток протоков.При раздражении чревных нервов, осуществляющих симпатическую иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет активации р-адренорецепторов медиатором норадреналином). Но торможение секреции сопровождается накоплением секреторного материала в панкреацитах. Поступление пищи в желудок во время приема пищи вызывает раздражение хемо- и механорецепторов желудка, что приводит к продолжению безусловно-рефлекторного сокоотделения поджелудочной железой, вызванного стимуляцией рецепторов ротовой полости.Поступление в двенадцатиперстную кишку желудочного содержимого вызывает наряду с дуоденопанкреатическим секреторным рефлексом (с рецепторов слизистой оболочки) выделение из эндокринных клеток гастроинтестинальных гормонов, стимулирующих или тормозящих сокоотделение поджелудочной железой. Роль основных гуморальных регуляторов выполняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усиливают влияние друг друга на панкреоциты, особенно на фоне выделения ацетилхолина в синаптических окончаниях холинергических нервных волокон, иннервирующих железу. Важным гуморальным возбудителем панкреатической секреции является гастрин, выделяемый У-клетками слизистой оболочки антрального отдела желудка. Возбуждающее влияние этих гормонов подкрепляется бомбезином, серотонином и инсулином. Тормозят выделение сока соматостатин, глюкагон, энкефалины, вещество Р, ТИП, ПП, а также кальцитонин и АКТГ. Но их роль в комплексной гуморальной регуляции поджелудочной железы изучена недостаточно. Секреция сока поджелудочной железой осуществляется в три фазы. Первая называется сложнорефлекторной. Она обусловлена комплексом условных и безусловных раздражителей, предшествующих и сопутствующих приему пищи. В первую фазу выделяется около 10—15 % от общего объема сока за три фазы, а ферментов — около 25 %. Во вторую желудочную фазу вырабатывается около 10 % сока от общего его объема (с высоким содержанием ферментов). В третью кишечную фазу выделяется основное количество сока (около 75 % от общего объема), но менее богатого ферментами. Его состав отличается большим содержанием бикарбонатов, необходимых для ощелачивания содержимого двенадцатиперстной кишки.
Билет 27
Функции мозжечка.
Мозжечок состоит из 2-х полушарий и червя между ними.
Серое вещество образует кору и ядра. Белое образовано отростками нейронов.
Мозжечок получает афферентные нервные импульсы от тактильных рецепторов, рецепторов вестибулярного аппарата, проприорецепторов мышц и сухожилий, а также двигательных зон коры. Эфферентные импульсы от мозжечка идут к красному ядру среднего мозга, ядру Дейтерса продолговатого мозга, к таламусу, а затем к моторным зонам КБП и подкорковым ядрам.Общей функцией мозжечка является регуляция позы и движений. Эту функцию он осуществляет путем координации активности других двигательных центров: вестибулярных ядер, красного ядра, пирамидных нейронов коры.
Двигательные функции:
1. Регуляцию мышечного тонуса и позы.
2. Коррекцию медленных целенаправленных движений в ходе их выполнения, а также координацию этих движений с рефлексам и положения тела.
3. Контроль за правильным выполнением быстрых движений, осуществляемых корой.
В связи с тем, что мозжечок выполняет данные функции, при его удалении развивается комплекс двигательных нарушений, называемый триадой Лючиани. Он включает:
1. Агония и дистония — снижение и неправильное распределение тонуса скелетных мышц.
2. Астазия — невозможность слитного сокращения мыши, сохранения устойчивого положения тела при стоянии, сидении (покачивание).
3. Астения — быстрая утомляемость мышц.
4. Атаксия — плохая координация движений при ходьбе. Неустойчивая «пьяная» походка.
5. Адиадохокинез — нарушение правильной последовательности быстрых целенаправленных движений.
В клинике умеренные поражения мозжечка проявляются триадой Шарко:
1. Нистагм глаз в состоянии покоя.
2. Тремор конечностей, возникающий при их движениях.
3.Дизартрия — нарушения речи.
Л. А.Орбели установил, что мозжечок влияет и на различные вегетативные функции, эти влияния могут, быть возбуждающими и тормозящими. Например, при раздражении мозжечка увеличивается или снижается кровяное давление, изменяется частота сердцебиений, дыхание, пищеварение. Мозжечок влияет на обмен веществ. Воздействует через вегетативные нервные центры, координируя их активность с движением. Ф-ии внутренних органов изменяются в связи с изменением обменных процессов в них. Поэтому мозжечок оказывает на них адаптационно-трофическое влияние.
Дата добавления: 2018-01-21; просмотров: 688; Опубликованный материал нарушает авторские права? | Защита персональных данных | ЗАКАЗАТЬ РАБОТУ
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Студент — человек, постоянно откладывающий неизбежность… 10457 — | 7301 — или читать все…
Источник
Секреция ПЖ находится под контролем нервных и гуморальных механизмов. Начальную секрецию ПЖ вызывают вид, запах пищи и другие раздражители (условнорефлекторные сигналы), а также жевание и глотание (безусловнорефлекторные сигналы). При этом нервные сигналы, формирующиеся в рецепторах полости рта и глотки, достигают продолговатого мозга, и затем эфферентные влияния по волокнам блуждающего нерва поступают к железе и вызывают её секрецию.
У человека с фистулой ГПП выделение панкреатического сока начинается через 2—3 мин после того, как он увидел пищу или услышат о ней. Это пример условнорефлекторного пути возбуждения панкреатической секреции. Симпатические волокна, иннервирующие ПЖ, тормозят её секреторную активность и в то же время изменяют реактивность железы по отношению к другим воздействиям, усиливая синтез органических веществ.
Торможение панкреатической секреции происходит при раздражении многих чувствительных нервов, при болевых реакциях, во время сна, при напряжённой физической и умственной работе.
Для стимуляции панкреатической секреции прямые нервные влияния имеют меньшее значение, нежели гуморальные. Ведущее значение в гуморальной регуляции секреции ПЖ принадлежит желудочно-кишечным гормонам.
Большинство регуляторов секреции ферментов ПЖ действуют на рецепторы мембраны ацинарных клеток, расположенные на их базолатеральной поверхности. Выделяют рецепторы ХК, бомбезина, ацетилхолина, субстанции Р, ВИП, секретина.
Стимуляторы панкреатической секреции. ВИП и секретин стимулируют панкреатическую секрецию, активируя аденилатциклазу. Как и в других типах клеток, аденилатциклаза способствует образованию циклического аденозинмонофосфата, в результате чего протеинкиназа А, усиливающая секрецию панкреатического сока, богатого бикарбонатами, переходит в активную форму. Другие стимуляторы поджелудочной секреции (ХК, ацетилхолин, гастрин-рилизинг пептид, субстанция Р) действуют на специфические рецепторы, во внутриклеточной передаче сигнала от которых задействованы альтернативные вторичные мессенджеры.
Эти вещества повышают внутриклеточное содержание циклического гуанозинмонофосфата, что приводит к увеличению внутриклеточного содержания инозитолтрифосфата, диацилглицерола, арахидоновой кислоты и кальция. Эти промежуточные вещества-посредники активируют различные протеинкиназы, в результате этого повышается секреция ферментов. Данные, полученные в опытах на животных, свидетельствуют, что действие комбинации агонистов на различные мембранные рецепторы может вызывать синергический, но не суммарный (аддитивный) эффект. Например, ХК увеличивает секрецию бикарбонатов, стимулированную секретином, но секретин не повышает секреторный ответ на действие ХК.
Ингибиторы секреции поджелудочной железы. Различные вещества, ответственные за подавление панкреатической секреции, действуют по принципу отрицательной обратной связи во время и после приёма пищи.
Панкреатический полипептид (ПП) представляет собой пептидный гормон, образующийся в островках Лангерганса и подавляющий панкреатическую секрецию воды, бикарбонатов и ферментов. Концентрация ПП в плазме крови возрастает после мнимого кормления, после приёма пищи, после экспериментального закисления среды ДПК, а также при стимуляции блуждающего нерва, при действии ХК, секретина, ВИП. ПП может выступать как антагонист мускариновых рецепторов и способен ингибировать выделение ацетилхолина из постганглионарных нейронов ПЖ; его конечный эффект проявляется на уровне ацинарных клеток.
Пептид YY высвобождается в дистальной части подвздошной кишки и и толстой кишке в ответ на поступление пищи смешанного характера, по жиры, находящиеся в просвете кишки, в большей степени способны стимулировать его секрецию. Пептид YY уменьшает чувствительность ПЖ к действию секретина и ХК, возможно, за счёт уменьшения секреции ацетилхолина и норадреналина и ингибирования выделения ХК слизистой оболочкой ДПК.
Соматостатин ингибирует секрецию секретина слизистой оболочкой ДПК, а также уменьшает чувствительность к секретину рецепторных полей. Единственный эффект соматостатина — снижение секреции ферментов и бикарбонатов ПЖ. Соматостатин синтезируют клетки слизистой оболочки желудка и кишечника, а также D-клетки островков Лангерганса. Его активность составляет только около 25% от активности гипоталамического соматостатина. Но только соматостатин, продуцируемый слизистой оболочкой тонкой кишки, оказывает угнетающее действие на секрецию ПЖ.
Выделение соматостатина происходит при участии автономной нервной системы в ответ на поступление жиров и аминокислот с пищей.
Соматостатин блокирует панкреатическую секрецию несколькими способами. Во-первых, он действует путём угнетения продукции стимулирующих пептидов (ХК). Во-вторых, посредством угнетающего воздействия на ЦНС и регулируя работу интрапанкреатических ганглиев, соматостатин ингибирует поступление ацетилхолина в пресинаптическую щель и, возможно, ингибирует выработку инсулина. Хотя рецепторы к соматостатину найдены также в ацинарных клетках, эти рецепторы скорее способствуют увеличению секреции, чем блокируют её.
Другие ингибиторы, представленные среди гормонов эндокринных клеток островков Лангерганса, включают панкреатический глюкагон и панкреастатин, а также нейропептиды: кальцнтонин-генерирующий пептид и энкефалины (табл. 1-3). Панкреатический глюкагон ингибирует секрецию ПЖ, стимулированную ХК, секретином или пищей; угнетает секрецию бикарбонатов, воды и ферментов. Панкреастатин ингибирует панкреатическую секрецию, тормозя высвобождение ацетилхолина эфферентными окончаниями блуждающего нерва. Кальцитонин-генерирующий пептид может проявлять свою активность через стимуляцию выделения соматостатина. Энкефалины и подобные им опиоиды снижают выделение секретина слизистой оболочкой ДПК и могут также ингибировать высвобождение ацетилхолина.
Таблица 1-3. Ингибиторные нейромодуляторы
Маев И.В., Кучерявый Ю.А.
Источник

