Островки поджелудочной железы развиваются из
ПЖ в эмбриогенезе формируется из двух зачатков кишечной энтодермы — дорсального и вентрального — и из мезенхимы. Дорсальный зачаток развивается раньше вентрального: он появляется на 3-й неделе эмбриогенеза из печёночного дивертикула и первичной кишечной трубки, в то время как вентральный — только на 4—5-й неделе из развивающейся ДПК (из протокового отрезка средней кишки).
Иногда появляются сразу два вентральных панкреатических зачатка. Возможностью образования двойного вентрального зачатка и последующего его аномального развития объясняется механизм формирования кольцевидной ПЖ, что рядом авторов рассматривается как атавизм.
Основные процессы дифференпировки морфологических структур ПЖ происходят в период с середины 6-й до 12-й недели внутриутробного развития. В середине 5-й недели гестации ПЖ и ДПК входят в состав единого мезенхимального комплекса, отграниченного от полости тела. Этот тканевой комплекс соединен вентральной брыжейкой с желудком, а дорсальной — с задней стенкой полости тела. Формирующаяся ПЖ на этой стадии развития представлена системой ветвящихся трубочек, расположенных в толще мезенхимы. На 5-й неделе эмбрионального развития уже определяются два изолированных друг от друга отдела ПЖ, один из которых (вентральная часть) располагается в едином тканевом комплексе с ДПК, а другой (дорсальная часть) — лежит свободно в толще дорсальной брыжейки.
На 6-й неделе эмбрионального развития вентральная часть ПЖ по-прежнему плотно прилежит к ДПК, располагаясь с ней в едином тканевом комплексе. В середине 7-й недели эмбрионального развития начинается слияние вентрального и дорсального отделов ПЖ. Вентральный зачаток при этом вращается по часовой стрелке позади ДПК и срастается с дорсальным зачатком.
После слияния панкреатических зачатков происходят последовательные изменения формы органа. Так, в зависимости от гестационного возраста эмбриона, ПЖ по форме напоминает запятую, затем имеет форму вопросительного знака, приобретает булавовидную, а затем S-образную форму. Начиная с 8-й недели развития железа напоминает «лежащую» латинскую букву «S».
С середины 11-й-начала 12-й недели впервые можно говорить о формировании крючковидного отростка, который огибает формирующиеся верхние брыжеечные сосуды. Из вентрального зачатка формируются крючковидный отросток и нижние 2/3 головки ПЖ, затем из дорсального зачатка развиваются шейка, тело, хвост и верхняя часть головки органа.
Из протока вентрального зачатка, срастающегося с дорсальным зачатком, развивается главный панкреатический (вирсунгов) проток (ГПП), который служит основным дренажом ПЖ (см. рис. 1-1, CD). Проксимальный проток дорсального зачатка, известный как добавочный (санториниев) проток ПЖ, обычно сохраняется и открывается в ДПК через малый сосочек.
Во 2-й половине внутриутробного развития появляются дольки ПЖ и соединительнотканные междольковые перегородки. В этот период определяется топография синтеза белков: цитоплазматическая локализация, а также увеличение темпов синтеза. На 6-м месяце в ациноцитах видны секреторные гранулы, содержащие белки с амило- и липолитической активностью. Дифференцировка клеток ацинусов и протоков завершается к концу беременности.
Во время роста и вращения ДПК (см. рис. 1-1, показано стрелкой, CD) вентральный зачаток ПЖ перемещается к дорсальному, и впоследствии они срастаются. Первоначально общий жёлчный проток прикрепляется к вентральной части ДПК и сдвигается вокруг её дорсальной части, в то время как сама ДПК вращается. ГПП формируется в результате сращения дистальной части дорсального панкреатического протока и всего вентрального панкреатического протока.

Рис.1-1.Эмбриогенез поджелудочной железы: а–г — схематическое изображение последовательных этапов развития поджелудочной железы с 5-й по 8-ю неделю эмбрионального развития; д–ж — схематические изображения поперечных срезов через двенадцатиперстную кишку и развивающуюся поджелудочную железу
Таким образом, к 12-й неделе внутриутробного развития в ПЖ определяются основные структурные образования в зачатковой форме, или на той или иной стадии формирования. Их последующая дифференцировка обеспечивает весь диапазон функционирования не только в пренатальном, но и в постнатальном онтогенезе. Эндокринные участки железы (островки) развиваются как зачатки из начальных отрезков переднего и заднего протоков между 10-й и 14-й неделями гестации. После 16-й недели эндокринная часть ПЖ отделяется от протоков, приобретает собственное кровоснабжение и становится независимой от протоковой системы железы.
Нервная и сосудистая системы ПЖ начинают формироваться на 6—7-й неделе внутриутробного развития.
Формирование кровеносной системы заканчивается одновременно с дифференцировкой апинарных клеток к 7-му месяцу гестации. Формирование иннервации заканчивается в постнатальном и детском периоде. Морфофункциональное развитие ПЖ завершается только в возрасте 18—20 лет. После 40—50-летнего возраста намечаются гипотрофические изменения органа, связанные со склеротической трансформацией его кровеносных сосудов.
Аномалии при повороте или сращении зачатков развивающейся ПЖ могут приводить к специфическим врождённым аномалиям, таким как агенезия (аплазия) ПЖ, удвоение и как частный его случай — расщеплённая ПЖ (pancreas divisum), эктопическая ткань селезёнки в ПЖ, кольцевидная (pancreas annulare), добавочная ПЖ (pancreas aberrans), аномальное, панкреатобилпарное соустье, холедохоцеле и др.
Маев И.В., Кучерявый Ю.А.
Опубликовал Константин Моканов
Источник
Островки Лангерга́нса — скопления гормон-продуцирующих (эндокринных) клеток, преимущественно в хвостовой части поджелудочной железы. Открыты в 1869 году немецким патологоанатомом Паулем Лангергансом (1849—1888). Островки составляют приблизительно 1—2 % массы поджелудочной железы. Поджелудочная железа взрослого здорового человека насчитывает около 1 миллиона островков (общей массой от одного до полутора граммов), которые объединяют понятием орган эндокринной системы.
Историческая справка[править | править код]
Пауль Лангерганс, будучи студентом-медиком, работая у Рудольфа Вирхова, в 1869 году описал скопления клеток в поджелудочной железе, отличавшиеся от окружающей ткани, названные впоследствии его именем[2][3]. В 1881 году К. П. Улезко-Строганова впервые указала на эндокринную роль этих клеток[4]. Инкреаторная функция поджелудочной железы была доказана в Страсбурге (Германия) в клинике крупнейшего диабетолога Наунина Mering и Minkowski в 1889 году — открыт панкреатический диабет и впервые доказана роль поджелудочной железы в его патогенезе[3]. Русский учёный Л. В. Соболев (1876—1919) в диссертации «К морфологии поджелудочной железы при перевязке её протока при диабете и некоторых других условиях» показал, что перевязка выводного протока поджелудочной железы приводит ацинозный (экзокринный) отдел к полной атрофии, тогда как панкреатические островки остаются нетронутыми. На основании опытов Л. В. Соболев пришёл к выводу: «функцией панкреатических островков является регуляция углеводного обмена в организме. Гибель панкреатических островков и выпадение этой функции вызывает болезненное состояние — сахарное мочеизнурение»[3].
В дальнейшем благодаря ряду исследований, проведенных физиологами и патофизиологами в различных странах (проведение панкреатэктомии, получение избирательного некроза бета-клеток поджелудочной железы химическим соединением аллоксаном), получены новые сведения об инкреаторной функции поджелудочной железы.
В 1907 году Lane & Bersley (Чикагский университет) показали различие между двумя видами островковых клеток, которые они назвали тип A (альфа-клетки) и тип B (бета-клетки).
В 1909 году бельгийский исследователь Ян де Мейер предложил называть продукт секреции бета-клеток островков Лангерганса инсулином (от лат. insula — островок). Однако прямых доказательств продукции гормона, влияющего на углеводный обмен, обнаружить не удавалось[3].
В 1921 году в лаборатории физиологии профессора J. Macleod в Торонтском университете молодому канадскому хирургу Фредерику Бантингу и его ассистенту студенту-медику Чарлзу Бесту удалось выделить инсулин.
В 1955 году Сангеру и соавторам (Кембридж) удалось определить последовательность аминокислот и строение молекулы инсулина[3].
В 1962 году Марлин и соавторы обнаружили, что водные экстракты поджелудочной железы способны повышать гликемию. Вещество, вызывающее гипергликемию, назвали «гипергликемическим-гликогенолитическим фактором». Это был глюкагон — один из основных физиологических антагонистов инсулина[3].
В 1967 году Донатану Стейнеру и соавторам (Чикагский университет) удалось обнаружить белок-предшественник инсулина — проинсулин. Они показали, что синтез инсулина бета клетками начинается с образования молекулы проинсулина, от которой в последующем по мере необходимости отщепляется С-пептид и молекула инсулина[3].
В 1973 году Джоном Энсиком (Вашингтонский университет), а также рядом учёных Америки и Европы была проведена работа по очистке и синтезу глюкагона и соматостатина[3].
В 1976 году Gudworth & Bottaggo открыли генетический дефект молекулы инсулина, обнаружив два типа гормона: нормальный и аномальный. последний является антагонистом по отношению к нормальному инсулину[3].
В 1979 году благодаря исследованиям Lacy & Kemp и соавторов появилась возможность пересадки отдельных островков и бета-клеток, удалось отделить островки от экзокринной части поджелудочной железы и осуществить трансплантацию в эксперименте. В 1979—1980 гг. при трансплантации бета-клеток преодолён видоспецифический барьер (клетки здоровых лабораторных животных имплантированы больным животным другого вида)[3].
В 1990 году впервые выполнена пересадка панкреатических островковых клеток больному сахарным диабетом[3].
Типы клеток[править | править код]
Данная диаграмма демонстрирует структурные отличия между панкреатическими островками крысы (вверху) и человека (внизу) pars ventralis pancreas (брюшная часть) — слева; pars dorsalis pancreas (спинная часть) — справа. Различные типы клеток окрашены по-разному: альфа-клетки — красным, бета-клетки — синим, дельта-клетки — фиолетовым, ПП-клетки — зелёным, эпсилон-клетки — жёлтым. Бета-клетки грызуна, в отличие от человека сгруппированы в характерное инсулиновое ядро.
Альфа-клетки[править | править код]
- Альфа-клетки составляют 15…20 % пула островковых клеток — секретируют глюкагон (естественный антагонист инсулина).
Бета-клетки[править | править код]
- Бета-клетки составляют 65…80 % пула островковых клеток — секретируют инсулин (с помощью белков-рецепторов проводит глюкозу внутрь клеток организма, активизирует синтез гликогена в печени и мышцах, угнетает глюконеогенез).
Дельта-клетки[править | править код]
- Дельта-клетки составляют 3…10 % пула островковых клеток — секретируют соматостатин (угнетает секрецию многих желез);
ПП-клетки[править | править код]
- ПП-клетки составляют 3…5 % пула островковых клеток — секретируют панкреатический полипептид (подавляет секрецию поджелудочной железы и стимулирует секрецию желудочного сока).
Эпсилон-клетки[править | править код]
- Эпсилон-клетки составляют <1 % пула островковых клеток — секретируют грелин[5][6] («гормон голода» — возбуждает аппетит).
Строение островка[править | править код]
Панкреатический островок является сложно устроенным функциональным микроорганом с определённым размером, формой и характерным распределением эндокринных клеток. Клеточная архитектура островка влияет на межклеточное соединение и паракринную регуляцию, синхронизирует высвобождение инсулина.
Долгое время считалось, что островки человека и экспериментальных животных сходны как по строению, так и по клеточному составу. Работы последнего десятилетия показали, что у взрослых людей преобладающим типом строения островков является мозаичный, при котором клетки всех типов перемешаны по всему островку, в отличие от грызунов, для которых характерен плащевой тип строения клеток, при котором бета-клетки формируют сердцевину, а альфа-клетки находятся на периферии. Однако, эндокринная часть поджелудочной железы имеет несколько типов организации: это могут быть единичные эндокринные клетки, их небольшие скопления, небольшие островки (диаметром < 100 мкм) и крупные (зрелые) островки.
Небольшие островки имеют у человека и грызунов одинаковое строение. Зрелые островки Лангерганса человека обладают выраженной упорядоченной структурой. В составе такого островка, окруженного соединительнотканной оболочкой, можно выявить дольки, ограниченные кровеносными капиллярами. Сердцевину долек составляет массив бета-клеток, на периферии долек в непосредственной близости с кровеносными капиллярами находятся альфа- и дельта-клетки. Таким образом, клеточная композиция островка зависит от его размера: относительное число альфа-клеток увеличивается вместе с размером островка, в то время как относительное число бета-клеток уменьшается[7].
См. также[править | править код]
Примечания[править | править код]
- ↑ 1 2 Foundational Model of Anatomy
- ↑ Langerhans P. Beiträge zur mikroskopischen Anatomie der Bauchspeicheldrüse : Inaugural-Dissertation, zur Erlangung der Doctorwürde in der Medicine und Chirurgie vorgelegt der Medicinischen Facultät der Friedrich-Wilhelms-Universität zu Berlin und öffentlich zu vertheidigen am 18. Februar 1869. — Berlin : Buchdruckerei von Gustav Lange, 1869.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 Клиническая диабетология / Ефимов А. С., Скробонская Н. А. — 1-е изд. — К.: Здоровья, 1998. — 320 с. — 3000 экз. — ISBN 5-311-00917-9.
- ↑ Жуковский М. А. Детская эндокринология. — 3-е изд. — М.: Медицина, 1995. — 656 с. — 8000 экз. — ISBN 5-225-01167-5.
- ↑ K. M. Andralojc, A. Mercalli, K. W. Nowak. et al. Ghrelin-producing epsilon cells in the developing and adult human pancreas (англ.).
- ↑ Epsilon cells (Cytokines&Cells Encyclopedia) (англ.). Архивировано 26 октября 2012 года.
- ↑ Прощина А. Е., Савельев С. В. Иммуногистохимическое исследование распределения α- и β-клеток в разных типах островков Лангерганса поджелудочной железы человека // Бюллетень экспериментальной биологии и медицины. — Изд. РАМН, 2013. — Т. 155, № 6. — С. 763—767.
Ссылки[править | править код]
Источник
Эмбриональное развитие поджелудочной железы
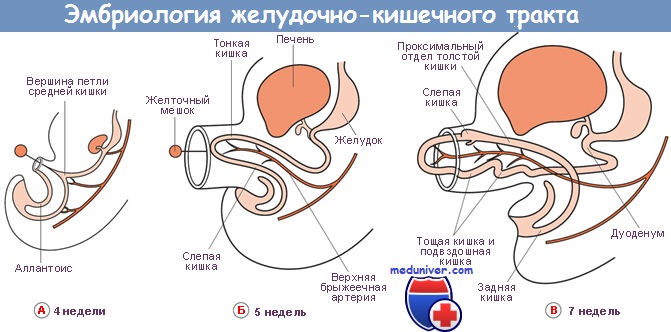
Поджелудочная железа развивается из отдельных зачатков — вентрального и дорсального, — образующихся из соединения эмбриональных передней кишки со средней. Дорсальный зачаток увеличивается влево и формирует основную массу зрелой железы.
Вентральный зачаток, тесно связанный с развивающимся общим желчным протоком, первоначально отходит от двенадцатиперстной кишки. Он соединяется с дорсальной системой только на седьмой неделе внутриутробного развития, после ротации позади эмбриональной двенадцатиперстной кишки, и расположен каудально от дорсального зачатка.
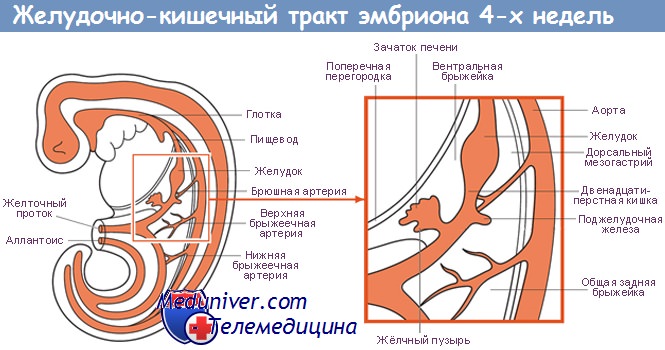
Обе части первичной поджелудочной железы содержат аксиальные протоки: дорсальный проток развивается из стенки двенадцатиперстной кишки, а вентральный — из общего желчного протока. При их слиянии вентральный проток (вирзунгов) продолжается дорсальным протоком (санториниев), образуя главный панкреатический проток.
Таким образом, общий желчный и панкреатический протоки открываются в двенадцатиперстную кишку на большом сосочке, тогда как часть дорсального протока внутри головки поджелудочной железы становится более или менее рудиментарной и открывается в двенадцатиперстную кишку проксимально от большого сосочка на небольшом добавочном или малом сосочке.
Полное отсутствие слияния систем двух протоков приводит к развитию разделенной поджелудочной железы. Такая аномалия встречается приблизительно у 5% людей и может быть предрасполагающим фактором панкреатита. Также возможно неполное слияние, при котором санториниев проток остается главным.
Недостаточный поворот тела вентрального зачатка по оси с устьем его протока может привести к развитию кольцевидной поджелудочной железы, окружающей вторую часть двенадцатиперстной кишки, что может вызвать обструкцию двенадцатиперстной кишки.
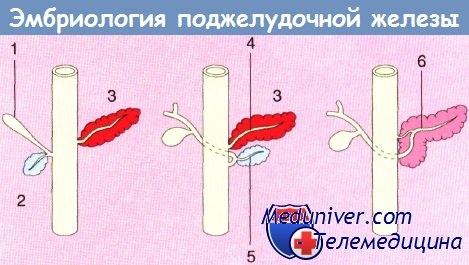
Схема, показывающая анатомию развивающейся поджелудочной железы:
1 — билиарный зачаток; 2 — вентральный;
3 — дорсальный; 4 — санториниев проток;
5 — вирзунгов проток; 6 — главный панкреатический проток.
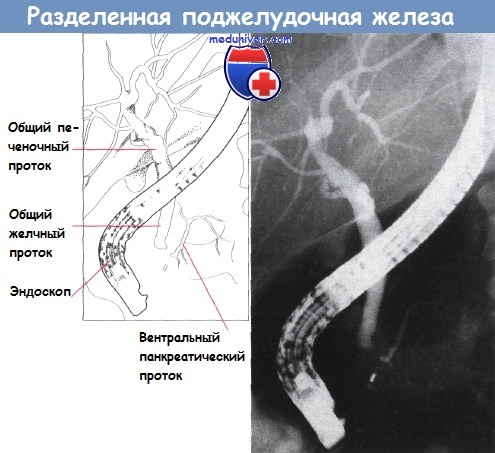
Разделенная поджелудочная железа. Определяются протоки в вентральной части железы.
Видно их отсутствие справа от позвоночника.
Хорошо заметно близкое расположение к поджелудочной железе общего желчного протока.
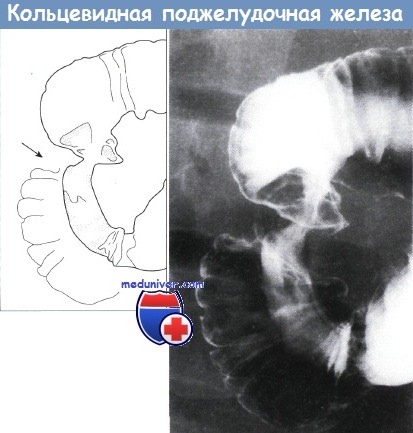
Верхний отдел желудочно-кишечного тракта, кольцевидная поджелудочная железа.
Наблюдается очаговое циркулярное ровное сужение середины второй части двенадцатиперстной кишки.
Дуоденальные складки и слизистая оболочка в остальном нормальные. Рентгенограмма с двойным контрастированием.
— Также рекомендуем «Острый панкреатит — диагностика»
Оглавление темы «Заболевания желудочно-кишечного тракта»:
- Сифилис заднего прохода — диагностика
- Гонорея прямой кишки — диагностика
- Предраковые состояния анальной области — диагностика
- Злокачественные опухоли анального канала — диагностика
- Врожденные аноректальные аномалии — диагностика
- Поджелудочная железа в норме — анатомия, методы обследования
- Эмбриональное развитие поджелудочной железы
- Острый панкреатит — диагностика
- Хронический панкреатит — диагностика
- Наследственный панкреатит — диагностика
Источник

