Островская часть поджелудочной железы
Эндокринная часть поджелудочной железы. Регенерация поджелудочной железы.
Островки состоят из эпителиальных клеток — панкреатических эндокриноцитов, или инсулоцитов. Величина островков, их форма и число входящих в состав клеток очень различны. Общее количество островков в поджелудочной железе достигает 1-2 млн. Средний размер островка 0,1-0,3 мм. Общий объем эндокринной части составляет около 3% всего объема железы. Островки пронизаны кровеносными капиллярами, окруженными перикапиллярным пространством. Эндотелий капилляров имеет фенестры, облегчающие поступление гормонов от инсулоцитов в кровь через перикапиллярное пространство.
В островковом эпителии различают 5 видов клеток: А-клетки, В-клетки, D-клетки, ВИП-клетки, РР-клетки.
А-клетки (альфа-клетки, или ацидофильные инсулоциты) — это крупные округлые клетки с бледным крупным ядром и цитоплазмой, содержащей ацидофильные гранулы. Гранулы обладают и аргирофилией. В состав этих гранул входит гормон глюкагон, расщепляющий гликоген и повышающий содержание сахара в крови.
А-клетки рассеяны по всему островку, образуя нередко небольшие скопления в центральной части. Они составляют около 20-25% от всех инсулоцитов.
В-клетки (бета-клетки, или базофильные инсулоциты) имеют кубическую или призматическую форму, крупное темное, богатое гетерохроматином ядро. Доля В-клеток достигает 70-75% от общего числа инсулоцитов. В цитоплазме В-клеток накапливаются осмиофильные гранулы, содержащие гормон инсулин. Инсулин регулирует синтез гликогена из глюкозы. При недостатке продукции инсулина глюкоза не превращается в гликоген, содержание ее в крови повышается и создаются условия для развития заболевания, называемого сахарным диабетом.
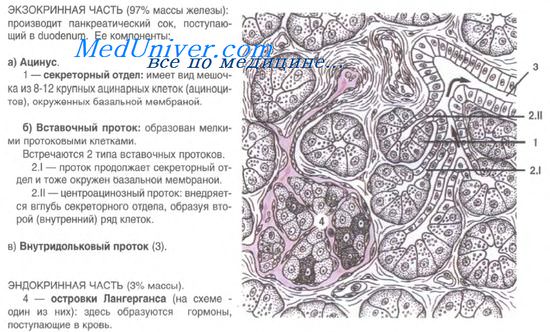
D-клетки (дельта-клетки, или дендритические инсулоциты) составляют 5-10% среди всех островковых клеток. Форма их иногда звездчатая с отростками. В цитоплазме определяются гранулы средних размеров и плотности. В гранулах накапливается гормон соматостатин. Он тормозит секрецию инсулина и глюкагона, снижает продукцию ряда гормонов желудочно-кишечного тракта — гастрина, секретина, энтероглюкагона, холецистокинина и др., подавляет секрецию соматотропного гормона в гипофизе.
ВИП-клетки (аргирофильные клетки) встречаются в островках в небольшом количестве. В цитоплазме выявляются плотные аргирофильные гранулы, содержащие вазоактиеный интестиналъный полипептид. Он обладает выраженным сосудорасширяющим дейтвием, снижает кровяное давление, угнетает секрецию соляной кислоты в желудке, стимулирует выделение глюкагона и инсулина.
РР-клетки — полигональной формы инсулоциты, расположенные преимущественно по периферии островка. Количество их — 2-5% от общего числа клеток островка. В цитоплазме РР-клеток выявляются мелкие гранулы, содержащие панкреатический полипептид. Основная роль панкреатического полипептида в организме — регуляция скорости и количества экзокринной секреции поджелудочной железы и желчи в печени. Таков клеточный состав островкового эпителия, представляющего собой мозаику дивергентно развивающихся клеточных дифферонов.
В дольках поджелудочной железы встречаются еще ацинозно-инсулярные клетки, в цитоплазме которых одновременно содержатся гранулы, характерные как для ацинозных, так и для островковых клеток.
Ткани поджелудочной железы иннервируют блуждающий и симпатический нервы. В интрамуральных вегетативных ганглиях находятся холинергические и пептидергические нейроны, волокна которых заканчиваются на клетках ацинусов и островков. Между нервными клетками ганглиев и островковыми клетками устанавливается тесная связь с образованием нейроинсулярных комплексов.
С возрастом в поджелудочной железе постепенно уменьшается количество островков. В островках наблюдаются закономерные возрастные изменения клеточных взаимоотношений, заключающиеся в быстрой смене после рождения преобладания А-клеток над В-клетками на преобладание В-клеток над А-клетками у взрослых. Затем происходит постепенное увеличение количества А-клеток, которое наряду с одновременным, хотя и незначительным, уменьшением числа В-клеток приводит нередко в пожилом и особенно старческом возрасте вновь к преобладанию А-клеток над В-клетками.
Регенерация поджелудочной железы. В эмбриогенезе островки растут благодаря пролиферации исходных клеток-предшественниц и их дивергентной дифференцировке в соответствующие клеточные диффероны. У взрослых физиологическая регенерация ацинозных и островковых клеток происходит в основном путем внутриклеточного обновления органелл. Митотическая активность клеток в связи с высокой специализацией низкая. После резекции части или повреждения органа наблюдается некоторое повышение уровня пролиферативной активности клеток ацинусов, протоков и островков, последующее образование новых ацинусов. Однако ведущей формой регенерации экзокринной части железы является регенерационная гипертрофия.
Восстановительные процессы в эндокринной части железы происходят за счет пролиферативной активности инсулоцитов и клеток эпителия протоков путем ацино-инсулярной трансформации.
— Также рекомендуем «Печень. Развитие печени. Строение печени.»
Оглавление темы «Строение желудка. Строение кишечника.»:
1. Пищевод. Слизистая пищевода. Строение стенки пищевода.
2. Желудок. Развитие желудка. Строение желудка. Железы желудка.
3. Эпителий желез желудка. Клетки желудка. Гормоны желудка.
4. Тонкая кишка. Развитие тонкой кишки. Строение тонкой кишки.
5. Эпителий тонкой кишки. Клетки тонкой кишки.
6. Толстая кишка. Развитие и строение толстой кишки. Червеобразный отросток.
7. Прямая кишка. Строение прямой кишки. Поджелудочная железа.
8. Эндокринная часть поджелудочной железы. Регенерация поджелудочной железы.
9. Печень. Развитие печени. Строение печени.
10. Гепатоциты. Строение гепатоцитов. Образование желчи.
Источник
В
эндокринной части паренхимы поджелудочной
железы располагаются островки
Лангерганса.
Их основными
структурными единицами являются
секреторные (α, β, Δ, F
и другие) клетки.
А-клетки
(α-клетки)
островков
продуцируют
глюкагон.
Он увеличивает гликогенолиз в печени,
снижает в ней утилизацию глюкозы, а
также повышает глюконеогенез и образование
кетоновых тел. Результатом этих
воздействий является увеличение
концентрации глюкозы в крови. Вне печени
глюкагон повышает липолиз и снижает
синтез белков.
На
-клетках
имеются рецепторы, которые при уменьшении
уровня глюкозы во внеклеточной среде
усиливают секрецию глюкагона. Секретин
угнетает продукцию глюкагона, а другие
желудочно-кишечные гормоны стимулируют
ее.
B-клетки
(-клетки)
синтезируют и накапливают инсулин.
Этот гормон
увеличивает проницаемость клеточных
мембран для глюкозы и аминокислот, а
также способствует превращению глюкозы
в гликоген, аминокислот в белки, а жирных
кислот в триглицериды.
Синтезирующие
инсулин клетки
способны реагировать на изменения
содержания в крови и просвете ЖКТ
калоригенных молекул (глюкозы, аминокислот
и жирных кислот). Из аминокислот наиболее
выражена стимуляция секреции инсулина
аргинином и лизином.
Поражение
островков Лангерганса приводит к гибели
животного из-за нехватки в организме
инсулина. Только
этот гормон снижает содержание глюкозы
в крови.
Д-клетки
(Δ-клетки)
островков синтезируют панкреатический
соматостатин.
В поджелудочной
железе он оказывает тормозящее
паракринное влияние на секрецию гормонов
островками Лангерганса (преобладает
влияние на -клетки),
а внешнесекреторным аппаратом —
бикарбонатов и ферментов.
Эндокринное
влияние панкреатического соматостатина
проявляется торможением секреторной
активности в ЖКТ, аденогипофизе,
паращитовидной железе и почках.
Наряду
с секрецией, панкреатический соматостатин
снижает сократительную активность
желчного пузыря и желчных протоков, а
на всем протяжении ЖКТ -уменьшает
кровообращение, моторику и всасывание.
Активность
Д-клеток возрастает привысоком
содержании в просвете пищеварительного
тракта аминокислот (особенно лейцина
и аргинина) и глюкозы, а также при
увеличении концентрации в крови ХКП,
гастрина, желудочного ингибирующего
полипептида (ЖИП) и секретина. В то же
время, норадреналин угнетает высвобождение
соматостатина.
Панкреатический
полипептидсинтезируется
F-клетками
(или РР-клетками) островков. Он
уменьшает
объем панкреатического секрета и
концентрацию в нем трипсиногена, а также
тормозит выведение желчи, но стимулирует
базальную секрецию желудочного сока.
Выработка
панкреатического полипептида стимулируется
парасимпатической нервной системой,
гастрином, секретином и ХКП, а также при
голодании, приеме богатого белками
корма, гипогликемии и физической
нагрузке.
Интенсивность
выработки гормонов поджелудочной железы
контролируется вегетативной нервной
системой (парасимпатические нервы
вызывают гипогликемию, а симпатические
— гипергликемию). Однако основными
факторами регуляции секреторной
активности клеток в островках Лангерганса,
являются концентрации питательных
веществ в крови и просвете ЖКТ. Благодаря
этому, своевременные реакции клеток
островкового аппарата обеспечивают
поддержание постоянного уровня
питательных веществ в крови между
приемами корма.
ЭНДОКРИННАЯ ФУНКЦИЯ ПОЛОВЫХ ЖЕЛЕЗ
После
наступления половой зрелости основными
источниками половых гормонов в организме
животных становятся постоянные половые
железы (у самцов — семенники, а у самок
— яичники). У самок периодически могут
появляться и временные эндокринные
железы (например, плацента во время
беременности).
Половые
гормоны делят на мужские (андрогены) и
женские (эстрогены).
Андрогены
(тестостерон,
андростендион, андростерон
и др.) специфически стимулируют рост,
развитие и функционирование органов
размножения самцов, а с наступлением
половой зрелости — образование и
созревание мужских половых клеток.
Еще
до рождения в организме плода формируются
вторичные половые признаки. Это в
значительной степени регулируется
образующимися в семенниках андрогенами
(секретируются клетками Лейдига) и
фактором, секретируемым клетками Сертоли
(находятся в
стенке семенного канальца).
Тестостерон обеспечивает дифференцировку
наружных половых органов по мужскому
типу, а секрет клеток Сертоли предотвращает
образование матки и маточных труб.
В
период полового созревания андрогены
ускоряют инволюцию тимуса, а
в других тканях стимулируют накопление
питательных веществ, синтез белка,
развитие мышечной и костной ткани,
повышают физическую работоспособность
и сопротивляемость организма
неблагоприятным воздействиям.
Андрогены
влияют на ЦНС (например, вызывают
проявления полового инстинкта). Поэтому
удаление половых желез (кастрация) у
самцов делает их спокойными и может
привести к нужным для хозяйственной
деятельности изменениям. Например,
кастрированные животные быстрее
откармливаются, мясо их вкуснее и нежнее.
До
рождения, секреция андрогенов
обеспечивается совместным действием
на плод ЛГ самки и хорионического
гонадотропина (ХГ). После рождения,
развитие семенных канальцев, спермиев
и сопровождающую эти процессы выработку
БАВ клетками Сертоли стимулирует
собственный гонадотропин самца — ФСГ,
а ЛГ вызывает
секрецию
тестостерона
клетками Лейдига.
Старение сопровождается угасанием
активности половых желез, но продолжается
выработка половых гормонов надпочечником.
К
видовым особенностям клеток Сертоли
семенников жеребца, быка и кабана
относится их способность кроме
тестостерона вырабатывать эстрогены,
которые регулируют обмен веществ в
половых клетках.
Яичники
в организме половозрелой самки в
соответствии со стадиями полового цикла
вырабатывают эстрогены
и гестагены.
Основным источником эстрогенов
(эстрона, эстрадиола и эстриола) являются
фолликулы, а гестагенов — желтое тело.
У
неполовозрелой самки эстрогены
надпочечников стимулируют развитие
репродуктивной системы (яйцеводов,
матки и влагалища) и вторичных половых
признаков (определенного телосложения,
молочных желез и т.д.). После наступления
половой зрелости, концентрация в крови
женских половых гормонов значительно
повышается за счет их интенсивной
выработки яичниками. Возникающие при
этом уровни эстрогенов
стимулируют
созревание половых клеток, синтез белков
и образование мышечной ткани в большинстве
внутренних органов самки, а также
повышают сопротивляемость ее организма
к вредным воздействиям и вызывают
связанные с половыми циклами изменения
в органах животного.
Высокие
концентрации эстрогена вызывают рост,
расширение просвета и усиление
сократительной активности яйцеводов.
В матке они повышают кровенаполнение,
стимулируют размножение клеток эндометрия
и развитие маточных желез, а также
изменяют чувствительность миометрия
к окситоцину.
У самок многих
видов животных эстрогены вызывают
ороговение клеток влагалищного эпителия
перед течкой. Поэтому качество гормональной
подготовки самки к спариванию и овуляции
выявляют по цитологическим анализам
вагинального мазка.
Эстрогены
также способствуют формированию
состояния «охоты» и соответствующих
половых рефлексов в наиболее благоприятную
для оплодотворения стадию полового
цикла.
После
овуляции, на месте бывшего фолликула
образуется желтое
тело.
Вырабатываемые им гормоны (гестагены)
влияют на матку, молочные железы и ЦНС.
Они вместе с эстрогенами регулируют
процессы зачатия, имплантации
оплодотворенной яйцеклетки, вынашивания
беременности, родов и лактации. Основным
представителем гестагенов является
прогестерон. Он стимулирует секреторную
активность маточных желез и делает
эндометрий способным реагировать на
механические и химические воздействия
разрастаниями, которые необходимы для
имплантации
оплодотворенной яйцеклетки
и образования плаценты. Прогестерон
также снижает
чувствительность матки к окситоцину и
расслабляет ее. Поэтому
преждевременное снижение концентрации
гестагенов в крови беременных самок
вызывает роды до полного созревания
плода.
Если
беременность не наступила, то желтое
тело подвергается инволюции (продукция
гестагенов прекращается) и начинается
новый овариальный цикл. Умеренные
количества прогестерона в синергизме
с гонадотропинами стимулируют овуляцию,
а большие — тормозят секрецию гонадотропинов
и овуляция не происходит. Небольшие
количества прогестерона также необходимы
для обеспечения течки и готовности к
спариванию. Кроме этого, прогестерон
участвует в формировании доминанты
беременности
(гестационной доминанты), направленной
на обеспечение развития будущего
потомства.
После
воздействия эстрогенов, прогестерон
способствует развитию железистой ткани
в молочной железе, что приводит к
формированию в ней секреторных долек
и альвеол.
Наряду
со стероидными гормонами желтое тело,
эндометрий и плацента, преимущественно
перед родами, продуцируют гормон
релаксин.
Его выработка стимулируется высокими
концентрациями ЛГ и вызывает повышение
эластичности лонного сочленения,
расслабление связки тазовых костей, а
непосредственно перед родами повышает
чувствительность миометрия к окситоцину
и вызывает расширению маточного зева.
Плацента
возникает в несколько этапов. Сначала,
в ходе дробления оплодотворенной
яйцеклетки образуется трофобласт.
После присоединения к нему внезародышевых
кровеносных сосудов трофобласт
превращается в хорион,
который после плотного соединения с
маткой становится сформировавшейся
плацентой.
У
млекопитающих плацента обеспечивает
прикрепление, иммунологическую защиту
и питание плода, выведение продуктов
обмена, а также выработку гормонов
(эндокринная функция), необходимых для
нормального течения беременности.
Уже
на ранних сроках беременности в местах
прикрепления ворсинок хориона к матке
вырабатываетсяхорионический
гонадотропин.
Его появление ускоряет развитие зародыша
и предотвращает инволюцию желтого тела.
Благодаря этому желтое тело поддерживает
высокий уровень прогестерона в крови
до тех пор, пока плацента сама не начнёт
синтезировать его в необходимом
количестве.
Вырабатываемые
в организме беременных самок негипофизарные
гонадотропины имеют видовые особенности,
но могут влиять на репродуктивные
функции и у других видов животных.
Например, введение гонадотропина
сыворотки крови жеребых кобыл (ГСЖК)
вызывает у многих млекопитающих выделение
прогестерона. Это сопровождается
удлинением полового цикла и задерживает
приход охоты. У коров и овец ГСЖК также
вызывает одновременный выход нескольких
зрелых яйцеклеток, что используется
при трансплантации эмбрионов.
Плацентарные
эстрогены
вырабатываются плацентой большинства
млекопитающих (у приматов — эстрон,
эстрадиол и
эстриол,
а у лошади — эквилин
и эквиленин)
преимущественно во второй половине
беременности из дегидроэпиандростерона
образующегося в надпочечниках плода.
Плацентарный
прогестерон
у ряда млекопитающих (приматы, хищники,
грызуны) секретируются в количествах
достаточных для нормального вынашивания
плода даже после удаления желтых тел.
Плацентарный
лактотропин
(плацентарный лактогенный гормон,
плацентарный пролактин,
хорионический соматомаммотропин)
поддерживает
рост плода, а у самки
увеличивает синтез белка в клетках и
концентрацию СЖК в крови, стимулирует
рост секреторных отделов молочных желёз
и их подготовку к лактации, а также
задерживает
в организме ионы кальция, снижает мочевую
экскрецию фосфора и калия.
По
мере увеличения сроков беременности в
крови самок растет уровень плацентарного
кортиколиберина,
который увеличивает чувствительность
миометрия к окситоцину. Данный либерин
практически не влияет на секрецию АКТГ.
Это связано с тем, что во время беременности
в крови растет содержание белка, который
быстро нейтрализует кортиколиберин и
он не успевает подействовать на
аденогипофиз.
ТИМУС
Тимус
(зобная или вилочковая железа) имеется
у всех позвоночных животных. У большинства
млекопитающих он состоит из двух
соединенных друг с другом долей,
расположенных в верхней части грудной
клетки сразу за грудиной. Однако, у
сумчатых животных эти доли тимуса обычно
остаются отдельными органами. У
пресмыкающихся и птиц железа обычно
имеет вид цепочек, расположенных по обе
стороны шеи.
Наибольших
размеров по отношению к массе тела тимус
большинства млекопитающих достигает
к моменту рождения. Затем он медленно
растет и в период полового созревания
достигает максимальной массы. У морских
свинок (и некоторых других видов животных)
крупный тимус сохраняется на протяжении
всей жизни, но у большинства высокоразвитых
животных после полового созревания
железа постепенно уменьшается
(физиологическая
инволюция), но
полной атрофии ее не происходит.
В
тимусе эпителиальные клетки продуцируют
тимические
гормоны влияющие эндокринным и паракринным
путем на гемопоэз, а также дифференцировку
и активность Т-клеток.
В
тимусе на предшественники Т-лимфоцитов
последовательно действуют тимопоэтин
и тимозины.
Они делают
дифференцирующиеся в тимусе клетки
чувствительными к активированному
кальцием тимулину
(или тимическому сывороточному фактору
— ТСФ).
П
р и м е ч а н и е: Возрастное снижение
содержания ионов кальция в организме
является причиной падения активности
тимулина у старых животных.
Секреторная
активность тимуса тесно связана с
деятельностью гипоталамуса и других
эндокринных желез (гипофиза, эпифиза,
надпочечников, щитовидной железы и
гонад). Гипоталамический
соматостатин, удаление
надпочечников и щитовидной железы
снижают выработку
тимических гормонов, а эпифиз
и кастрация усиливают гормонопоэз в
тимусе.
Кортикостероиды
регулируют распределение тимических
гормонов между тимусом, селезенкой и
лимфоузлами, а тимэктомия приводит к
гипертрофии коры надпочечников.
Перечисленные
примеры свидетельствуют о том, что
вилочковая железа обеспечивает интеграцию
нейро-эндокринной и иммунной систем в
целостном макроорганизме.
ЭПИФИЗ
Эпифиз
(шишковидная железа) расположена у
позвоночных под кожей головы или в
глубине мозга. Основными клетками
эпифиза у млекопитающих являются
пинеалоциты,
а у более
примитивных животных здесь имеются и
фоторецепторы. Поэтому,
наряду с эндокринной функцией эпифиз
может обеспечивать ощущение степени
освещенности объектов. Это позволяет
глубоководным рыбам осуществлять
вертикальную миграцию в зависимости
от смены дня и ночи, а миногам и
пресмыкающимся — оберегать себя от
опасности сверху. У
некоторых перелетных птиц эпифиз,
вероятно, выполняет функцию навигационных
приборов при перелетах.
Эпифиз
земноводных уже способен вырабатывать
гормон мелатонин,
которыйуменьшение
количество пигмента в клетках кожи.
Пинеалоциты
непрерывно синтезируют гормон серотонин,
который в темное время суток и при низкой
активности симпатической нервной
системы (у птиц и млекопитающих)
превращается в мелатонин. Поэтому
продолжительность дня и ночи, влияют
на содержание этих гормонов в эпифизе.
Возникающие при этом ритмические
изменения их концентрации в шишковидной
железе определяют у животных суточный
(циркадианный) биологический ритм
(например, периодичность сна и колебания
температуры тела), а также влияет на
формирование таких сезонных реакций
как зимняя спячка, миграция, линька и
размножение.
Увеличение
содержания мелатонина в эпифизе оказывает
снотворный, анальгезирующий и седативный
эффекты, а также тормозит
половое созревание молодняка.
Поэтому после удаления эпифиза у цыплят
быстрее наступает половое созревание,
у самцов млекопитающих — гипертрофируются
семенники и усиливается созревание
спермиев, а у самок — удлиняется период
жизни желтых тел и увеличивается матка.
Мелатонин
снижает секрецию ЛГ, ФСГ, пролактина и
окситоцина. Поэтому низкий уровень
мелатонина в светлое время суток
способствует усилению молокообразования
и высокой половой активности животных
в те времена года, когда ночи наиболее
короткие (весной и летом). Мелатонин
также нейтрализует повреждающее действие
стрессоров и является естественным
антиоксидантом.
У
млекопитающих серотонин и мелатонин
выполняют свои функции в основном в
эпифизе, а дистантными гормонами железы,
вероятно, являются полипептиды.
Значительная
их часть наряду с кровью, секретируется
в спинномозговую жидкость и через нее
поступает в различные отделы ЦНС. Это
оказывает преимущественно тормозное
влияние на поведение животного и другие
функции мозга.
В
эпифизе уже обнаружено
около 40 секретирующихся в кровь и
спиномозговую жидкость биологически
активных пептидов. Из них наиболее
изучены антигипоталамические факторы
и адреногломерулотропин.
Антигипоталамические
факторы обеспечивают связь эпифиза с
гипоталамо-гипофизарной системой. К
ним, например, относятся аргинин-вазотоцин
(регулирует секрецию пролактина) иантигонадотропин(ослабляет секрецию
ЛГ).
Адреногломерулотропин
стимулируя выработку альдостерона
надпочечником, влияет на водно-солевой
обмен.
Таким
образом, основной
функцией эпифиза
является регуляция
и координация биоритмов.
Посредством контроля деятельности
нервной и эндокринной систем животного,
шишковидная железа обеспечивает
опережающую реакцию его систем на смену
времени суток и сезона.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
