Топография поджелудочной железы у свиньи
Поджелудочная железа (Pancreas) — железа двойной функции: внешнесекреторной и внутрисекреторной. Внешнесекреторная функция заключается в синтезе и выделении в двенадцатиперстную кишку сока, содержащего пищеварительные ферменты и электролиты, внутрисекреторная — в синтезе и выделении в кровь гормонов.
Внешнесекреторная часть железы сильно развита и составляет более 95 % ее массы. Она имеет дольчатое строение и состоит из альвеол (ацинусов) и выводных протоков. Основная масса ацинусов (железисто-пузырьковидные концевые отделы) представлена панкреатическими клетками — панкреацитами — секретируемыми клетками.
Внутрисекреторная часть железы представлена островками Лангерганса, которые составляют около 30 % массы железы. Различают несколько видов островков Лангерганса по способности секретировать полипептидные гормоны: А-клетки продуцируют глюкогон, В-клетки — инсулин, D-клетки — самостатин. Основную массу островков Лангерганса (около 60 %) составляют В-клетки.
Поджелудочная железа лежит в брыжейке двенадцатиперстной кишки, на печени, разделяясь на правую, левую и среднюю доли. Проток поджелудочной железы открывается в двенадцатиперстную кишку самостоятельно или вместе с желчным протоком. Иногда встречается добавочный проток, который впадает в двенадцатиперстную кишку самостоятельно. Иннервируется поджелудочная железа симпатическими и парасимпатическими нервами (n. vagus).
У собак железа длинная, узкая, красноватого цвета, образует более объемистую левую ветвь и более длинную правую ветвь, достигающую почек. Поджелудочный проток открывается в двенадцатиперстную кишку вместе с желчным протоком. Иногда встречается добавочный проток. Абсолютная масса железы 13-18 г.
У крупного рогатого скота поджелудочная железа располагается вдоль двенадцатиперстной кишки от 12-го грудного до 2-4-го поясничного позвонка, под правой ножкой диафрагмы, частично на лабиринте ободочной кишки. Состоит из поперечной и правой продольной ветвей, соединяющихся под углом в правой стороне. Выводной проток открывается обособленно от желчного протока на расстоянии 30-40 см от него (у овец вместе с желчным протоком). Абсолютная масса железы у крупного рогатого скота 350-500 г, у овец 50-70 г.
У лошадей на поджелудочной железе различают среднюю часть — тело, прилежащее к воротному изгибу двенадцатиперстной кишки. Левый конец железы, или хвост, длинный и узкий, достигает слева слепого мешка желудка, соединяясь с ним, селезенкой и левой почкой. Правый конец железы, или головка, доходит до правой почки, слепой и ободочной кишки. Поджелудочный проток открывается вместе с печеночным. Иногда встречается дополнительный проток. Цвет железы желтоватый, абсолютная масса до 250-350 г.
У свиней на железе различают среднюю, правую и левую доли. Через среднюю долю проходит воротная вена печени. Железа лежит под двумя последними грудными и двумя первыми поясничными позвонками. Проток один, открывается на 13-20 см дистальнее устья желчного протока. Абсолютная масса железы 150 г.
Внешнесекреторная (экзогенная) функция поджелудочной железы. Основной продукт внешнесекреторной функции поджелудочной железы — пищеварительный сок, который содержит 90 % воды и 10% плотного осадка. Плотность сока 1,008-1,010; рН 7,2-8,0 (у лошадей 7,30-7,58; у крупного рогатого скота 8). В состав плотного осадка входят белковые вещества и минеральные соединения: бикарбонат натрия, хлорид натрия, хлорид кальция, фосфорнокислый натрий и др.
Сок поджелудочной железы содержит протеолитические и нуклеолитические ферменты (трипсин, хемотрипсин, карбоксипептидазы, эластазу, нуклеазы, аминопептидазу, коллагеназу, дипептидазу), амилолитические ферменты (а-амилазу, мальтазу, лактазу, инвертазу) и липолитические ферменты (липазу, фосфолипазу, холинэстеразу, карбоксиэстеразу, моноглицеридлипазу, щелочную фосфатазу). Трипсин расщепляет белки до аминокислот и выделяется в виде неактивного трипсиногена, который активируется ферментом кишечного сока энтерокиназой. Химотрипсин расщепляет белки и полипептиды до аминокислот и выделяется в форме неактивного химотрипсиногена; активируется трипсином. Карбоксиполипептидазы действуют на полипептиды, отщепляя от них аминокислоты. Дипептидазы расщепляют дипептиды на свободные аминокислоты. Эластаза действует на белки соединительной ткани — эластин, коллаген. Протаминаза расщепляет протамины, нуклеазы — нуклеиновые кислоты на мононуклеотиды и фосфорную кислоту.
При воспалении поджелудочной железы, аутоиммунных процессах протеолитические ферменты становятся активными уже в самой железе, вызывая ее разрушение. а-Амилаза расщепляет крахмал и гликоген до мальтозы; мальтаза — мальтозу до глюкозы; лактаза расщепляет молочный сахар на глюкозу и галактозу (она имеет существенное значение в пищеварении молодняка), инвертаза — сахарозу на глюкозу и фруктозу; липаза и другие липолитические ферменты расщепляют жиры на глицерин и жирные кислоты. Липолитические ферменты, в частности липаза, секретируются в активном состоянии, но расщепляют только жир, эмульгированный желчными кислотами. Амилазы, также как и липазы, в соке поджелудочной железы находятся в активном состоянии.
Из электролитов в соке поджелудочной железы содержатся натрий, калий, хлор, кальций, магний, цинк, медь и значительное количество бикарбонатов, обеспечивающих нейтрализацию кислого содержимого двенадцатиперстной кишки. Тем самым создается оптимальная среда для активных ферментов.
Доказано, что помимо перечисленного выше действия сок поджелудочной железы обладает свойством регуляции микробной ассоциации в двенадцатиперстной кишке, оказывая определенное бактерицидное действие. Прекращение поступления в кишечник панкреатического сока ведет к усиленному бактериальному росту в проксимальном отделе тонкого кишечника у собак.
Эндокринная (гормональная) функция поджелудочной железы. Важнейшими гормонами поджелудочной железы являются инсулин, глюкогон и соматостатин.
Инсулин образуется в В-клетках из предшественника — проинсулина. Синтезируемый проинсулин поступает в аппарат Гольджи, где расщепляется на молекулу С-пептида и молекулу инсулина. Из аппарата Гольджи (пластинчатый комплекс) инсулин, С-пептид и частично проинсулин поступают в везикулы, где инсулин связывается с цинком и депонируется в таком состоянии. Под влиянием различных стимулов инсулин освобождается от цинка и поступает в прекапиллярное пространство. Основным стимулятором секреции инсулина служит глюкоза: при ее повышении в крови усиливается синтез инсулина. В определенной степени этим свойством обладают аминокислоты аргинин и лейцин, а также глюкогон, глетрин, секретин, глюкокортикоиды, соматостатин, никотиновая кислота. Инсулин в крови находится в свободном и связанном с белками плазмы состоянии. Распад инсулина происходит в печени под влиянием глютатионтрансферазы и глютатионредуктазы, в почках под влиянием инсулиназы, в жировой ткани под влиянием протеолитических ферментов. Проинсулин и С-пептид тоже подвергаются дегидратации в печени. Его биологическое действие обусловлено способностью связываться со специфическими рецепторами клеточной цитоплазматической мембраны.
Инсулин усиливает синтез углеводов, белков, нуклеиновых кислот и жира. Он ускоряет транспорт глюкозы в клетки инсулинозависимых тканей (печень, мышцы, жировая ткань), стимулирует синтез гликогена в печени и подавляет глюконеогенез (образование глюкозы из неуглеводных компонентов), гликогенолиз (распад гликогена), что в конечном итоге ведет к снижению уровня сахара в крови. Этот гормон ускоряет транспорт аминокислот через цитоплазматическую мембрану клеток, стимулирует синтез белка. Инсулин участвует в процессе включения жирных кислот в триглицериды жировой ткани, стимулирует синтез липидов и подавляет липолиз (распад жира).
В регуляции синтеза белка и утилизации углеводов вместе с инсулином участвуют кальций и магний. Концентрация инсулина в крови человека 15-20 мкЕД/мл.
Глюкогон — полипептид, секреция которого регулируется глюкозой, аминокислотами, гастроинтестинальными гормонами (панклеозимин) и симпатической нервной системой. Секреция глюкогона усиливается при снижении в крови сахара, СЖК, раздражении симпатической нервной системы, а угнетается при гипергликемии, повышении уровня СЖК, соматостатина. Под влиянием глюкогона стимулируется глюконеогенез, ускоряется распад гликогена, т. е. увеличивается продукция глюкозы. Под действием глюкогона ускоряется синтез активной формы фосфорилазы, участвующей в образовании глюкозы из неуглеводных компонентов (глюконеогенез). Глюкогон способен связываться с рецепторами адипацитов (клеток жировой ткани), способствуя распаду триглицеридов с образованием глицерина и СЖК. Глюконеогенез сопровождается не только образованием глюкозы, но и промежуточных продуктов обмена веществ — кетоновых тел, развитием кетоацидоза. Содержание в плазме крови глюкогона у человека составляет 50-70пг/мл. Концентрация этого гормона в крови увеличивается при голодании (голодный кетоз у овец), хронических заболеваниях печени.
Соматостатин — гормон, основной синтез которого осуществляется в гипоталамусе, а также в D-клетках поджелудочной железы. Соматостатин подавляет секрецию СТГ, АКТГ, ТТГ, гастрина, глюкогона, инсулина, ренина, секретина, вазоактивного желудочного пептида, желудочного сока, панкреатических ферментов и электролитов. Содержание соматостатина в крови повышается при сахарном диабете I типа, D-клеточной опухоли поджелудочной железы (соматостатиноме). Говоря о гормонах поджелудочной железы, следует отметить, что энергетический баланс в организме поддерживается сплошными биохимическими процессами, в которых непосредственное участие принимают инсулин, глюкогон и частично соматостатин. Так, во время голодания уровень в крови инсулина снижается, а глюкогона повышается, усиливается глюконеогенез. Благодаря этому поддерживается минимальный уровень глюкозы в крови. Усиление липолиза сопровождается повышением в крови СЖК, которые используются сердечной и другими мышцами, печенью, почками в качестве энергетического материала. В условиях гипогликемии источником энергии становятся и кетокислоты.
Нейроэндокринная регуляция функции поджелудочной железы. Деятельность поджелудочной железы находится под влиянием парасимпатической (n. vagus) и симпатической (чревные нервы) нервной системы, гипоталамогипофизарной системы и других желез внутренней секреции. В частности, блуждающий нерв играет определенную роль в регуляции ферментообразования. Секреторные волокна входят также в состав симпатических нервов, иннервирующих поджелудочную железу. При стимуляции отдельных волокон блуждающего нерва с усилением сокоотделения происходит и его торможение. Основоположником отечественной физиологии И. П. Павловым доказано, что отделение поджелудочного сока начинается при виде корма или раздражении рецепторов полости рта и глотки. Этот феномен необходимо учитывать в случаях назначения голодной диеты при остром панкреатите у собак, кошек и других животных, не допуская их зрительного и обонятельного контакта с кормом.
Наряду с нервной происходит и гуморальная регуляция функции поджелудочной железы. Поступление соляной кислоты в двенадцатиперстную кишку вызывает секрецию поджелудочного сока даже после перерезки блуждающих и чревных (симпатических) нервов и разрушения продолговатого мозга. Это положение лежит в основе назначения медикаментов, снижающих секрецию поджелудочного сока при остром панкреатите. Под влиянием соляной кислоты желудочного сока, поступающего в кишечник, из клеток слизистой оболочки тонкой кишки выделяется просекретин. Соляная кислота активирует просекретин, превращая его в секретин. Всасываясь в кровь, секретин действует на поджелудочную железу, усиливая выделение ею сока: одновременно он тормозит функцию обкладочных желез, чем препятствует чрезмерно интенсивной секреции соляной кислоты железами желудка. Секретин в физиологическом отношении является гормоном. Под влиянием секретина образуется большое количество поджелудочного сока, бедного ферментами и богатого щелочами. Учитывая эту физиологическую особенность, лечение острого панкреатита направлено на снижение секреции соляной кислоты в желудке, подавление активности секретина.
В слизистой оболочке двенадцатиперстной кишки образуется также гормон панкреозимин, который усиливает образование ферментов в поджелудочном соке. Подобное действие оказывают гастрин (образуется в желудке), инсулин, соли желчных кислот.
Тормозящее влияние на секрецию панкреатического сока оказывают нейропептиды — гастроингибирующий полипептид (ГИП), панкреатический полипептид (ПП), вазоактивный интерстинальный полипептид (ВИП), а также гормон соматостатин.
При лечении плотоядных животных с нарушением внешнесекреторной функции поджелудочной железы необходимо иметь в виду, что на молоко выделяется мало сока, на мясо, черный хлеб — много. При кормлении мясом выделяется много трипсина, при кормлении молоком — много липазы и трипсина.
Источник
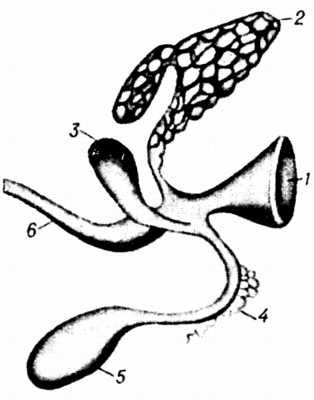
Рис. 1. Развитие печени и поджелудочной железы.
Рис. 1. Развитие печени и поджелудочной железы:
1 желудок;
2 дорзальный зачаток поджелудочной железы;
3 вентральный зачаток поджелудочной железы;
4 зачаток печени;
5 зачаток жёлчного пузыря;
6 зачаток двенадцатиперстной кишки
(по Гертвигу с изменениями).
поджелу́дочная железа́ (Pancreas), пищеварительная железа с внешне- и внутрисекреторными функциями, расположенная в брыжейке двенадцатиперстной кишки, на печени.
Эмбриогенез и анатомия. П. ж. (рис. 1) развивается из дорзального и вентрального выростов двенадцатиперстной кишки. В ней образуются 2 выводных протока главный и добавочный. Первый происходит из вентрального зачатка, второй из дорзального. В постэмбриональный период остаётся один главный проток, который впадает в двенадцатиперстную кишку вместе с жёлчным протоком. У лошади и часто у собаки сохраняются оба протока, а у крупного рогатого скота остаётся только добавочный. П. ж. у крупного рогатого скота (рис. 2) располагается в правом подреберье, занимая область от 12-го ребра до 34-го поясничного позвонка, и состоит из правой и левой ветвей. Выводной проток выходит в области правой ветви и впадает в двенадцатиперстную кишку на расстоянии 3040 см от впадения жёлчного протока (у овец вместе с жёлчным протоком). У свиньи различают среднюю, правую и левую доли П. ж. (рис. 3). П. ж. лежит под двумя последними грудными и двумя первыми поясничными позвонками. Среднюю долю прободает воротная вена печени. Главный выводной проток впадает в двенадцатиперстную кишку вместе с жёлчным протоком. У лошади П. ж. состоит из тела, доходящего до изгиба двенадцатиперстной кишки, левой части (или хвоста), достигающей слепого мешка желудка и связанной с ним, а также с селезёнкой и левой почкой соединительной тканью, и толстой правой части (или головки), доходящей до правой почки, слепой и ободочной кишок и прилегающей к ножкам диафрагмы.
Гистология.
П. ж. сложная трубчато-альвеолярная железа. Покрыта тонкой соединительнотканной капсулой, от которой внутрь органа отходят перегородки, разделяющие его на дольки. Вместе с перегородками в П. ж. проходят, разветвляясь, кровеносные сосуды и выводные протоки. Эпителий протоков, вначале однослойный призматический, по мере ветвления протоков становится однослойным кубическим. Самые мелкие ветви выводных протоков проникают внутрь долек и называются вставочными отделами. Их эпителийоднослойный, почти плоский. Заканчивается проток расширением (альвеолой ацинусом), состоящим из более высоких железистых клеток. Внутри альвеолы расположены клетки вставочного отдела, называемые центроацинозными. Между структурами (экзокринная часть П. ж.), описанными выше, имеются пронизанные сетями капилляров скопления эпителиальных клеток панкреатические (Лангерганса) островки (рис. 4). Они не связаны с выводными протоками и являются эндокринной частью П. ж. Островки развиваются из клеток вставочных отделов, могут образовываться даже у взрослых животных, состоят из а-клеток (тип А), б-клеток (тип Б) и клеток типа Д.
Физиология. П. ж. главная пищеварительная железа. Её паренхима (альвеолы и системы выводных протоков) осуществляет образование и выделение в двенадцатиперстную кишку панкреатического сока (экзокринная функция). Последний содержит ферменты (трипсиноген, трипсин, химотрипсин, карбоксилаза и эластаза, дезоксирибонуклеаза, а также рибонуклеаза, нуклеаза, амилаза и др.), необходимые для переваривания всех пищевых веществ. Островки Лангерганса не связаны с выводными протоками и выделяют свои продукты гормоны непосредственно в кровь (эндокринная функция П. ж.). Клетки типа А синтезируют глюкагон, повышающий содержание глюкозы в крови, клетки типа Б инсулин, способствующий накоплению гликогена в печени. Эпителий мелких выводных протоков образует липотропное вещество липокаин, препятствующий жировому перерождению печени.
Патология. Заболевания П. ж. вызываются неправильным кормлением, микробами, гельминтами, физическими факторами, поражением паращитовидной железы. Эти причины приводят к воспалению П. ж. (панкреатит), что ведёт к нарушению пищеварения и других функций. Значительное поражение островков Лангерганса приводит к развитию диабета сахарного, гипергликемии, гипогликемии.
Литература:
Иванов И. Ф., Ковальский П. А., Цитология, гистология, эмбриология, 3 изд., М., 1976.
См. также литературу при статье Пищеварение.
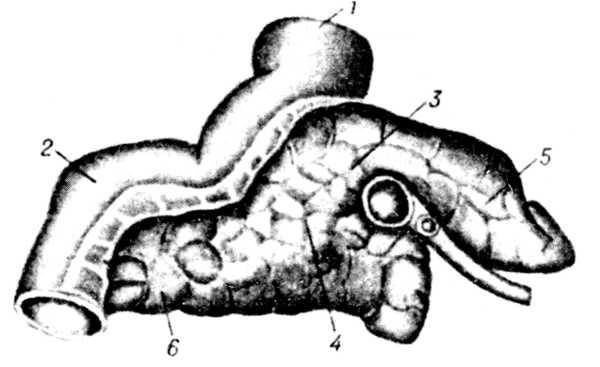
Рис. 2. Поджелудочная железа крупного рогатого скота.
Рис. 2. Поджелудочная железа крупного рогатого скота:
1 пилорус;
2 двенадцатиперстная кишка;
3 поджелудочная железа;
4 тело железы;
5 левая доля;
6 правая доля
(по Коху).
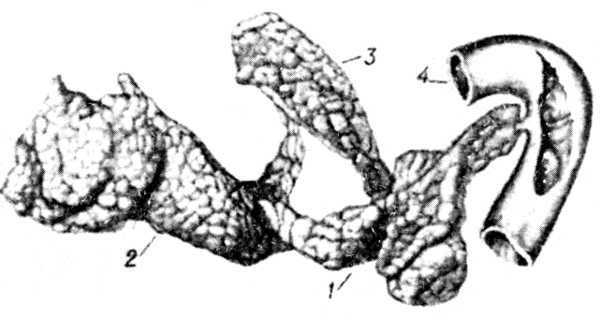
Рис. 3. Поджелудочная железа свиньи.
Рис. 3. Поджелудочная железа свиньи:
1 средняя доля;
2 левая доля;
3 правая доля;
4 отверстие воротной вены
(по Никкелю).
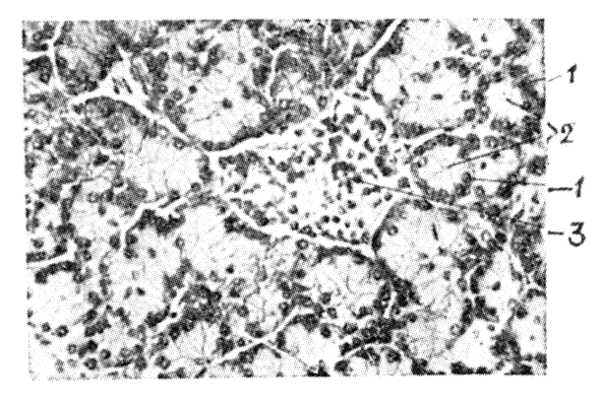
Рис. 4. Гистологическое строение поджелудочной железы.
Рис. 4. Гистологическое строение поджелудочной железы:
1 периферический участок клеток железы;
2 центральный участок клеток железы;
3 панкреатический островок.
Ветеринарный энциклопедический словарь. — М.: «Советская Энциклопедия».
Главный редактор В.П. Шишков.
1981.
Источник
Содержание к диссертации
Введение
1. Обзор литературы 10
1.1. Особенности развития, топографии и макро-микроскопического строения псджелудочной железы животных и человека 10
1.2. Артериальное русло поджелудочной железы животных и человека 18
1.3. Микроциркулягорное русло подаелудочной железы животных и человека 23
1.4. Венозное русло поджелудочной железы животных и человека 26
1.5. Лимфатическое русло поджелудочной железы животных и человека 29
2. Собственные исследования 39
2.1. Материал и методика исследования 39
2.2. Возрастные особенности топографии и строения сосудов поджелудочной железы свиньи 45
2.2.1. Особенности топографии и морфометрическая характеристика массы и линейных размеров поджелудочной железы свиньи 45
2.2.2. Особенности артериальных сосудов поджелудочной железы свиньи 64
2.2.2.1. Экстраорганные артериальные сосуды 64
2.2.2.2. Интраорганные артериальные сосуды 83
2.2.2.3. Артериальное звено микроциркуляторного русла.. 93
2.2.3. Особенности венозных сосудов поджелудочной железы свиньи 108
2.2.3.1, Интраорганные венозные сосуды 108
2.2.3.2. Экстраорганные венозные сосуды 112
2.2.4. Особенности лимфатического русла поджелудочной железы свиньи 121
2.2.4.1. Интраорганное лимфатическое русло 121
2.2.4.2. Архитектоника и морфометрия лимфангионов экстраорганных лимфатических сосудов поджелудочной железы 130
2.2.4.3. Регионарные лимфатические узлы поджелудочной железы 136
2.2.4.4. Изучение моторики лимфатических сосудов поджелудочной железы 140
3. Обсувдение результатов исследования 148
Выводы и практические предложения 159
Список исполъзованной литературы 163
- Артериальное русло поджелудочной железы животных и человека
- Микроциркулягорное русло подаелудочной железы животных и человека
- Лимфатическое русло поджелудочной железы животных и человека
- Возрастные особенности топографии и строения сосудов поджелудочной железы свиньи
Артериальное русло поджелудочной железы животных и человека
Поджелудочная железа млекопитающих имеет богатую васкуляриза-цию, что необходимо для нормального функционирования органа в обмене веществ. Ю.Г.Бойко (1970) подчеркивает, что сосудистым нарушениям принадлежит решающее значение в возникновении тканевых изменений поджелудочной железы. Поэтому изучение сосудистой системы органа в норме и при патологии приобретает большое значение.
Сосуды, осуществляющие кровообмен в поджелудочной железе всеядных, описаны недостаточно полно. Кроме коротких сведений в руководствах по анатомии домашних животных (Д.М.Автократов, 1949; А.И.&саевский, С.М.Боголюбский, В.Н.Жеденов, М.И.Лебедев, 1965), нам встретилось незначительное число источников, в которых описываются сосуды, принимающие участие в кровоснабжении этого органа у свиней.
Ю.П.Антипчук (1959) описал некоторые особенности роста и развития сосудов желудочно-кишечного тракта у свиней с 21 дня утробного развития и до шестимесячного возраста. Он выяснил, что в период с полутора- и до двухмесячного возраста у плодов свиньи заверпается, в основном, формирование экстраорганной системы желудка и кишечника. В эмбриональный период, указывает автор, относительный прирост площади сечения просвета артериальных сосудов пи щеварительного канала у свиней всегда намного превышает рост вен. В постэмбриональном периоде увеличение просвета вен идет быстрее артерий, что связано, по мнению автора, с началом активной после-утробной деятельности желудочно-кишечного тракта. Отмечено снижение интенсивности васкуляризации пищеварительного канала у свиней с возрастом. Сравнивая влияние различных типов кормления на сосудистую систему желудка и кишечника, автор отмечает, что концент-ратный тип кормления с повышенным уровнем белка животного происхождения ведет к образованию более тонких и мелкопетлистых капиллярных сосудистых сетей слизистой оболочки желудка и кишечника свиньи, свойственных плотоядным животным, по сравнению с полукон-центратным типом кормления.
Данные по интенсивности васкуляризации пищеварительного тракта можно отнести, по-видимому, и к сосудам поджелудочной железы свиней, хотя в своей работе Ю.П.Антипчук не остановился детально на этом вопросе.
Несколько позднее появилась работа 1.А.іуеевой (I96I) по источникам артериального кровоснабжения поджелудочной железы свиней. Автор отмечает, что питание этого органа происходит от ветвей чревной и краниальной брыжеечной артерий и, в редких случаях,толь-ко от чревной артерии. В работе приводятся также сведения о кровоснабжении каждой доли железы, но не указано число ветвей и их мор-фометрические данные. Левая доля железы, пишет автор, васкуляри-зуется за счет артериального сосуда, отходящего от селезеночной артерии, который вступает в начало доли, проходит в ней на всем протяжении и ветвится по магистральному типу. Головка железы по данным Л.А.іусевой, получает питание от селезеночной, печеночной, желудочно-двенадцатиперстной передней подаелудочно-двенадцатиперстной артерий. Они перед вступлением в головку делятся на два сосуда, которые вступают в нее с противоположных сторон и образуют передние и задние ветви, анастомозирующие между собой. Тип ветвления внутриоргандах сосудов этой доли рассыпной и смешанный. Правая доля железы получает артериальные сосуды преимущественно за счет краниальной брыжеечной артерии, редко — чревной артерии. Отмечено, что чаще всего сосуды, отходящие от печеночной, желудоч-но-двенадцатиперстной и передней поджелудочно-двенадцатиперстной артерий, располагаются поперек железы, а ветви селезеночной артерии — вдоль ее левой доли.
На основании проведенных исследований автор приходит к выводу, что внутриорганше ветви в поджелудочной железе свиней ана-стомозируют между собой и эти анастомозы образуют коллатерали внутреннего порядка.
Кровоснабжение поджелудочной железы у других видов домашних, а также лабораторных животных и человека в онтогенезе изучено подробнее, чем у всеядных животных.
У крупного рогатого скота установлены источники артериального кровоснабжения и пути оттока венозной крови от органа (Д.М.Автократов, 1949; П.В.Груздев, 1970; П.В.Груздев, С.И.Маланчук, 1980). Авторы, описывая васкуляризацию желудка и кишечника, касаются вопроса кровоснабжения поджелудочной железы, но приводят лишь схематичное описание артерий и вен этого органа.
Микроциркулягорное русло подаелудочной железы животных и человека
Советскими и зарубежными морфологами ( R.chambers, Zwei-fach B.W., 1944; Barcroft, Dornhost, 1954; Г.И.Мчедлишвили,1958; С.Н.Касаткин, I960; В.В.Куприянов, 1969; В.В.Куприянов, В.И.Козлов, I97I; Б.Фолков, Э.Нил, 1976; Я.Л.Караганов, В.В.Куприянов, В.И.Козлов, 1975; Я.Л.Караганов, Н.В.Кердиваренко, В.И.Левин, 1982; К.А.Шошенко, А.С.Голуб, В.Й.Брод, 1982) изучены особенности микроциркуляторного русла многих органов у человека и животных. Авторами приводятся четыре классификации капиллярного русла, основанные на особенностях в нем кровотока, в связи с внешней формой, электронно-микроскопическому строению стенки и особенностям транспорта веществ через стенку капилляра. Они взаимно дополняют друг друга, показывая тем самым многообразие признаков, характеризующих морфологию микроциркуляторного русла.
Для поджелудочной железы характерным является фрагаентация микроциркуляторного звена, которая обеспечивает известную самостоятельность каждого фрагмента системы. И.Ф.Крутикова (1970) указывает на возможность изолированных перестроек сосудистого русла в пределах отдельных претерминальшх долек.
Кровоснабжение каждой дольки осуществляется двумя-тремя доль-ковыми артериями, отходящими от артериального сплетения, расположенного в междольковых соединительнотканных прослойках. По мнению ряда авторов (Н.С.Шадрина, 1967; Н.М.Тотоев, 1969; В.Ф.Вдовин, А.А.Буравин, 1976; Н.А.Малков, I98I), в экскреторной паренхиме и в островках Дангерганса долек железы человека и животных располагается пространственная трехмерная сеть кровеносных капилляров.
Начальная часть артериального русла железы представлена капиллярами концевых отделов экскреторной паренхимы, инкреторной части, соединительнотканной стромы и выводных протоков.
Н.М.Тотоевым (1969) установлены возрастные отличия сети кровеносных капилляров экскреторной паренхимы долек поджелудочной железы человека. Они выражаются преимущественно в ее количественных изменениях и находятся в соответствии с перестройкой внутреннего строения органа в разные периоды жизни. Наиболее густые сети кровеносных капилляров отмечаются у детей первых лет жизни и подростков. С возрастом происходит постепенное разряжение капиллярных сетей: петли их увеличиваются и становятся более полиморфными,охватывая по два-три концевых отдела.
Среди исследователей нет единого мнения о кровоснабжении ин креторной части железы. Как считает Н.С.Шадрина (1967), васкуля-ризация островкового аппарата может происходить как непосредственно от междольковых артерий, так и от капилляров ацинозних отделов, которые получают кровь от ветвей междольковых артерий. Автор приходит к выводу, что кровеносное русло эндокринной и экзокринной части не обособлено друг от друга, а капилляры ацинусов мельче капилляров островков. Изучая сосуды инкреторного аппарата, Н.С.Шадрина обнаружила, что пронизывая островок между тяжами островковых клеток в различных направлениях, капилляры сплетаются в сосудистый клубочек, что подтверждает исследования И.Ф.Крутиковой,проведенные в 1970 году.
В 1969 году Н.М.Тотоев опубликовал данные, полученные при изучении строения и кровоснабжения островков Лангерганса поджелудочной железы человека в возрастном аспекте. Он отмечает наличие одного или нескольких «приносящих» сосудов, которые образуют сосудистый клубочек, и более тонкого выносящего сосуда. Он так же как и Н.С.Шадрина (1967) наблюдал многочисленные соединения капилляров клубочков с капиллярами экскреторной паренхимы.
В.Ф.Парфентьева и Ф.В.Морару (1976) также убеждены в том,что у плодов человека, как и у взрослых, в островках Лангерганса имеется сеть извитых широких капилляров. В.Ф.Ддовин (1976) согласен с теми авторами, которые утверждают, что каждый панкреатический островок имеет «приносящую» артериолу, отходящую от внутридолько-вой или междольковой артерий. В стенке такого сосуда обнаружены гладкомышечные клетки. Капиллярные клубочки панкреатических островков образованы округлыми и полигональными петлями без определенной ориентации, которые связаны с ацинарной паренхимой многочисленными сосудами.
Лимфатическое русло поджелудочной железы животных и человека
Внутриорганное лимфатическое русло поджелудочной железы животных изучено крайне недостаточно. Сведения в руководствах по анатомии домашних животных касаются в основном топографии регионарных лимфатических узлов, в которые происходит отток лимфы от органа. Данных по строению лимфатического русла поджелудочной железы свиньи в литературе мы не нашли.
Более подробно система лимфоциркуляции изучена на поджелудочной железе лабораторных животных и человека. Первое специальное исследование лимфатической системы поджелудочной железы провели P.Hoggan и G.Hoggan (1881) на трупах грызунов (кролики и мыши). Используя метод серебрения, авторы обнаружили на поверхности концевых камер собирающие лимфатические сосуды в виде петель и слепых расширений, которые сливались с выносящими лимфатическими сосудами. Рядом с кровеносными проходили периваскулярные лимфатические сосуды и попарно соединялись, увеличиваясь в диаметре. Но несмотря на большой калибр, по мнению авторов, они не являются главными путями оттока лимфы. E.ciein (1882) считал, что внутри-органная лимфатическая система поджелудочной железы человека представлена сосудами, не оформленными эндотелием, или открыто начинающимися периальвеолярными пространствами в паренхиме органа. Автор отметил, что эти пространства, соединяясь друг с другом,переходят в междольковые и продолжаются вдоль артерий, вен и протоков в виде лимфатических сосудов с эндотелиальными стенками и клапанами. По наблюдениям E.ciein периваскулярные лимфатические сосуды и капилляры идут параллельно и часто анастомозируют.
P.Sappey (1885) применил для изучения лимфатических сосудов поджелудочной железы метод внутритканевой инъекции и в противоположность P.Hoggan, G.Hoggan, E.Clein (1882) обнаружил,ЧТО KOp-нями лимфатической системы органа являются лимфатические капилляры, расположенные не на поверхности концевого отдела, а вокруг долек в соединительной ткани. Он заметил, что отток лимфы от железы идет в четырех направлениях (вверх, вниз, вправо, влево).
Наиболее полно описал лимфатическое русло поджелудочной железы собаки P.Bartels (1904). Он нашел лимфатические коллекторы и регионарные узлы, отметил тесную связь поджелудочной железы с лимфатическими сосудами и узлами желчного пузыря, желудка, двенадцатиперстной кишки, поперечной ободочной кишки, брыжейки тонкого отдела кишечника, левого надпочечника и почки. P.Sappey (1885) и P.Bartels (1904) указывали на то, чтО расположение лимфатических сосудов может объяснить переход воспалительных процессов с органов брюшной полости на поджелудочную железу лимфагенным путем.
На препаратах поджелудочной железы трупов 8 собак и 19 новорожденных и взрослых людей И.И.Косицын (I94I) обнаружил, что началом лимфатической системы органа является сеть лимфатических капилляров. Начинающиеся от них отводящие лимфатические сосуды охватывают дольки петлями и сопровождают кровеносные стволы и протоки. Автор описал локализацию регионарных лимфатических узлов поджелудочной железы и рассмотрел направление тока лимфы от разных частей поджелудочной железы.
Исследуя направление отводящих лимфатических сосудов и локализацию регионарных у лов на железе 90 трупов плодов и детей,Б.И. брагян (1955) выделил зоны железы, соответствующие ее эмбриональным зачаткам и источникам кровоснабжения (печеночную, верхнюю брыжеечную и селезеночную зоны) и определил группы регионарных у лов для них. Автор считал, что лимфатические капилляры имеют замкнутые начала в паренхиме органа и образуют пространственную сеть. Они вливаются в междольковые лимфатические сосуды, окружающие дольки железы, и формируют между ними крупнопетлистую сеть, которая анастомозирует с сетеобразными лимфатическими футлярами кровеносных сосудов и протоков.
Д.А.Ждановым (1955, 1958) установлено, что в поджелудочной железе человека трехмерная сеть лимфатических капилляров вставлена в сплетение отводящих лимфатических сосудов. Заложенные в соединительнотканных оболочках этих долек или долей лимфатические капилляры впадают в сосуды сплетения. Автором обнаружено, что междольковые лимфатические сосуды вливаются в сплетения более крупных лимфатических сосудов второго или третьего порядка, главным образом на поверхности поджелудочной железы.
Возрастные особенности топографии и строения сосудов поджелудочной железы свиньи
Изучение топографии поджелудочной железы свиньи, ее взаимоотношения с другими органами проводилось методом описания,зарисовки и препарирования на трупах животных и уточнялось при разделке туш на мясокомбинате.
В результате исследования десяти плодов в возрасте 90-110 дней и 32 животных с 15-дневного возраста и до двух лет установлены возрастные и индивидуальные особенности расположения поджелудочной железы (Pancreas).
На поджелудочной железе различают дорсальную (facies dorsa-lis) и вентральную поверхность (f.ventraiis). Дорсальной поверхностью орган прилегает к вентральным мышцам позвоночного столба в области последних двух грудных и первых двух поясничных позвонков. Вентральной поверхностью поджелудочная железа обращена к органам брюшной полости (рис.1).
Как с дорсальной, так и с вентральной поверхности орган покрыт листками брюшины, представленными брыжейкой двенадцатиперстной кишки.
У всех исследованных животных орган имеет три доли: правую (lobus dexter), среднюю или телО (lobus media) И левую долю (lobus sinister) (рис.2).
Поджелудочная железа имеет четыре края: краниальный и каудальниЙ (margo cranialis et caudalis), правый И левый (margo dexter et sinister). Краниальный край расположен над малой кривизной желудка, в области пилоруса и дивертикула, тесно прилегая к нему. Каудальный край поджелудочной железы приподнят дорсо-каудально и в области средней доли граничит с двенадцатиперстной кишкой. Правый и левый края органа приподняты в каудальном направлении. Правый край прилегает к начальному отрезку двенадцатиперстной кишки, а левый граничит с дивертикулом желудка и селезенкой,
В названии отделов поджелудочной железы свиньи (долей, поверхностей, краев) мы исходим из отношения их к продольной оси тела животного в соответствии с Международной ветеринарной анатомической номенклатурой (1979).
Правая доля расположена вдоль краниальной части двенадцатиперстной кишки. Начинаясь о
